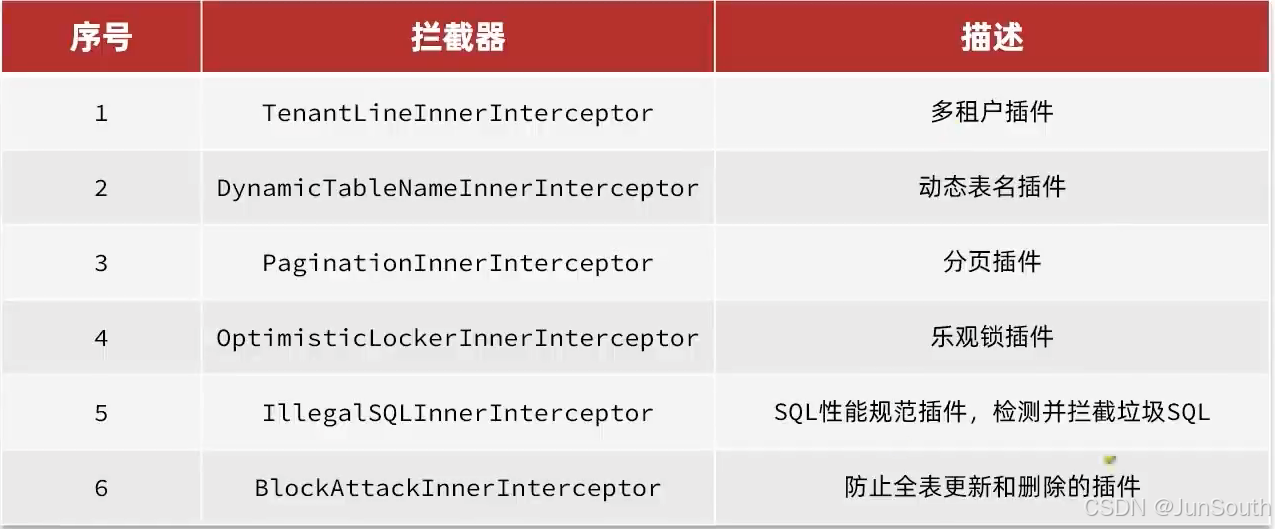
Engineering the Substrate Specificity of UDP-Glycosyltransferases for Synthesizing Triterpenoid Glycosides with a Linear Trisaccharide as Aided by Ancestral Sequence Reconstruction
通过祖先序列重建辅助工程化UDP-糖基转移酶的底物特异性,用于合成具有线性三糖的三萜糖苷
祖先重组图的推断与应用-文献精读59

摘要
三萜类化合物在医药和农业领域有广泛的应用。通过UDP-糖基转移酶(UGTs)催化三萜类化合物的糖基化是一种重要方法,用于生产具有增强功能的有价值衍生物。然而,已报道的仅有少数UDP-葡萄糖基转移酶能够在C3-OH位合成带有线性三糖的稀有三萜类化合物。本研究发现,UGT91H亚家族主要促进三萜类化合物的2"-O糖基化,且具有很高的区域选择性,随后通过祖先序列重建(ASR)进一步扩展了其底物范围。以祖先酶UGT91H_A1为模型,研究了序列-结构-功能关系。RTAS环(R212/T213/A214/S215)被鉴定为影响UGT91H_A1底物特异性的关键因素。将该RTAS环转移至其他UGT91H酶的对应位置,成功扩展了它们的底物谱。分子动力学模拟和量子力学计算进一步阐明了RTAS环的功能作用。UGT91H_A1被应用于结合自给式UDP-鼠李糖再生系统的低成本合成带有线性三糖的萜类鼠李糖苷。最后,我们开发了一个基于系统发生学的平台,用于高效地从植物基因组数据中挖掘新的UGT91H酶。本研究提供了用于合成带有线性三糖的多种三萜糖苷的强大生物催化剂,并展示了ASR作为工程化UDP-糖基转移酶功能的有效工具。
引言
三萜类化合物是一类重要的植物天然产物,已有超过2000种被报道。【1】由于其优良的生物活性,三萜类广泛应用于制药和农业领域。【2】然而,三萜类化合物的疏水性碳骨架导致其水溶性低,从而限制了其在临床医学中的生物利用度和治疗应用。【3】糖基化是对三萜类化合物进行结构修饰的关键方法,通过引入各种糖基,不仅丰富了它们的分子库,还显著提高了它们的水溶性和药理活性。【4】三糖修饰(包括线性和支链三糖)是三萜类化合物糖基化的重要类型,对于提高其药理活性具有重要意义。例如,具有线性三糖的戟皂苷I、戟皂苷H和通脱皂苷A表现出对多种肿瘤细胞的抑制活性。【5】
化学合成和酶促合成是天然产物糖基化的主要方法。化学合成面临多重问题,包括复杂的反应条件、有毒的反应试剂以及手性异构体的分离。【6】相比之下,酶促合成作为一种高效且环保的策略,受到特别关注。尤其是在天然产物的复杂多糖修饰中,酶的特异性区域选择性和对映选择性相较于化学合成中的多步保护-脱保护要求具有显著优势。【7】UDP-糖基转移酶(UGTs)是天然产物糖基化的主要酶类,它们利用各种UDP-糖(如UDP-Glc、UDP-Rha、UDP-Xyl、UDP-GlcA和UDP-Gal)作为糖基供体。【8】迄今为止,随着植物基因组学研究的发展,已经发现并表征了大量UGTs。【9】根据UGT命名委员会的统计数据(最后更新:2023-08-14)(https://labs.wsu.edu/ugt),约有9400种UGTs被分类和命名,其中约2000种UGTs能够催化三萜糖苷的合成,但大多数仅能催化三萜类化合物苷元上的单糖修饰。【9a,10】能够生成三萜多糖苷的UGTs极为稀少,限制了复杂三萜糖苷的酶促合成。目前,已有一些UGTs被报道能够催化三萜类化合物的单糖基团糖基化。【11】由于对极高的区域选择性的要求,只有少数UGTs能够催化三萜类化合物的二糖链延长。其中,大豆(Glycine max)的UGT91H4,甘草(Glycyrrhiza uralensis)的UGT91H9和UGT91H11可以作为2"-O-糖基转移酶,将糖基转移至三萜糖苷二糖链的第二个糖基,从而生成带有线性三糖的三萜糖苷。【12】因此,有必要挖掘更多强大的UGTs,用于合成多样化的带有线性三糖的三萜糖苷。然而,受限于缺乏高效的酶挖掘策略,这一进展受到限制。
蛋白质工程是获取用于合成三萜类三糖苷的高效UGTs的另一种途径。然而,这些UGTs的底物识别机制尚不清楚,且缺乏基于合理设计或半合理设计的关键位点突变指导。相比之下,祖先序列重建(ASR)作为一种日益成熟的方法,可以利用现存酶的系统发育关系推导祖先酶的序列,从而揭示酶家族的进化历史,并获得与新型或增强的酶学特性相关的特定序列。【13】最近的研究表明,重建的祖先酶不仅表现出更高的活性、热稳定性和更广泛的底物范围,【14】还可作为阐明现存酶催化机制的有力工具。【15】然而,ASR在生成高效UGTs方面的应用仍未得到充分探索。
在本研究中,我们系统地表征了七种针对三萜糖苷的UGT91H,并通过基于ASR构建祖先酶进一步扩展了其底物范围。随后,我们鉴定出一个RTAS环(R212/T213/A214/S215)影响UGT91H酶的底物特异性。通过分子动力学(MD)模拟和量子力学(QM)计算进一步阐明了RTAS环的功能作用。UGT91H亚家族的强大祖先酶UGT91H_A1成功应用于结合自给式UDP-鼠李糖再生系统的低成本合成具有线性三糖的萜类鼠李糖苷。最后,我们开发了一个基于系统发育的高效平台,用于从植物基因组数据中挖掘新的UGT91H酶。本研究提供了用于合成多种带有线性三糖的三萜糖苷的强大生物催化剂,并展示了ASR作为工程化UGTs底物特异性的有效工具的潜力。
结果与讨论
能够合成带有线性三糖的三萜糖苷的UGT91H酶

UGT91H4、UGT91H9和UGT91H11已被报道能够催化三萜糖苷在C2"-OH位的糖基化反应。【12】其中,UGT91H11被证实可作为2"-O-鼠李糖基转移酶,催化33种三萜双糖苷的鼠李糖基化反应。UGT91H4和UGT91H9仅对大豆皂苷Ⅲ和大豆皂苷Ⅳ显示活性。因此,我们首先探索了UGT91H4和UGT91H9的底物范围。由于UGT91H9在大肠杆菌中表现出较差的可溶性表达,我们重点验证了UGT91H4的催化特性。通过镍-亚氨基二乙酸亲和层析纯化后(图S1),首先以五环三萜GA 3-O-β-D-葡萄糖基-(1→2)-β-D-葡萄糖醛酸苷(GA 3-O-GlcA-Glc, 1)为受体,UDP-鼠李糖作为糖基供体,表征了UGT91H4的功能(图1和图S2-S9)。在保留时间3.1分钟处出现了一个新峰,被命名为产物1a。液相色谱-串联质谱(LC-MS/MS)分析表明,产物1a的[M-H]^-离子比底物1高146 Da,确认了UGT91H4能够将一个鼠李糖基转移到底物1上。通过放大酶促反应获得纯化的1a,并通过NMR光谱确认了其分子结构。C-2"/H-1'''的HMBC相关性表明,1a中引入的鼠李糖基连接在C2"-OH上。随后优化了反应条件,UGT91H4在pH 7.5和35 ℃时表现出最大活性(图S10)。接着,我们进一步测试了UGT91H4对另一受体GA 3-O-β-D-半乳糖基-(1→2)-β-D-葡萄糖醛酸苷(GA 3-O-GlcA-Gal, 2)的催化特性,UDP-鼠李糖作为糖基供体。通过LC-MS/MS和NMR分析获得了高纯度的反应产物(图S11-S18)。结果表明,UGT91H4能够特异性催化2的C2"-OH处的鼠李糖基化反应。因此,UGT91H4可作为2"-O-鼠李糖基转移酶,具有高度的区域特异性,类似于UGT91H11。
考虑到同一亚家族的酶由于其高度的序列和结构同源性,通常表现出相似的功能。【16】我们从NCBI数据库(www.ncbi.nlm.nih.gov, 表S1)中收集了六个新发现的UGT91H酶,它们的功能尚不明确。所有这六种UGT91H酶在大肠杆菌中均成功以可溶形式表达(图S1)。随后,使用底物1和2以及UDP-鼠李糖对这六种UGT91H酶进行了表征。根据这些六种UGT91H酶催化产物的新峰与UGT91H4催化的1a和2a的保留时间一致性(图S2和S11),我们推测UGT91Hs主要作为针对三萜糖苷的2"-O-糖基转移酶。我们随后研究了UGT91Hs对糖基受体的底物特异性,并探讨了底物结构对酶活性的影响。测试了30种具有不同苷元骨架和糖链的结构多样的三萜化合物(图2)。其中,底物1-22在C3-OH处带有二糖链,底物23-24、26-27在C3-OH处带有单糖基团,底物25、28-30没有糖基修饰。此外,底物1-22根据C3-OH上二糖链还原端的糖基分为四组:葡萄糖醛酸基团(GlcA, 1-7)、葡萄糖基团(Glc, 8-15)、阿拉伯糖基团(Ara, 16-18)和岩藻糖基团(Fuc, 19-22)。LC-MS/MS分析表明,七种UGT91H酶能够催化14种底物(1-5, 7-15)的鼠李糖基化反应。每个产物的[M-H]^-离子与其理论分子量一致,并显示出比底物高146 Da(图S19-S30)。根据反应结果,这些鼠李糖基化反应仅发生在C3-OH处含有二糖链且还原端为GlcA或Glc的底物上。没有糖基或仅带有单糖基团的底物没有检测到糖基化活性。
为了进一步确认UGT91Hs对四环三萜(8-14)的催化特性,我们选择底物9作为代表底物,并制备了由UGT91H8催化生成的产物9a进行NMR表征(图S31-S37)。C-2"/H-1'''的HMBC相关性表明9a中引入的鼠李糖基连接在C2"-OH上。随后,我们还验证了所有七种UGT91H酶对具有与8-14相同骨架且仅在R1位点带有二糖基团的绞股蓝皂苷XVII没有活性,表明UGT91Hs不能将糖基转移到四环三萜骨架的R1位点(图S38)。这些结果排除了糖基连接在8-14的R1位点,并进一步验证了我们之前的结论,即UGT91Hs作为四环和五环三萜的2"-O-糖基转移酶发挥作用。
从表1所示的反应转化中,我们发现了几条关于酶-底物关系的普遍规则,这些规则可以作为设计适合UGT91Hs底物的参考。(i)C3-OH位点二糖链上的第二个糖基类型影响UGT91Hs的催化效率。例如,底物1、2和3在二糖链上的第二个糖基分别为葡萄糖(Glc)、半乳糖基(Gal)和葡萄糖醛酸基(GlcA)。UGT91Hs对底物2表现出最高的催化活性,其次是底物1,最后是底物3。这表明UGT91Hs更倾向于催化具有Gal或Glc作为二糖链上第二个糖基的底物。(ii)带有C3-OH位点以外糖基的三萜类化合物更容易被UGT91Hs催化。具体而言,在几个具有相同三萜骨架和C3-OH二糖链的底物中(如底物4和5、6和7、10和11、12、13、14),底物5比底物4在C28-COOH位点多一个糖基,底物7比底物6在C28-COOH位点多一个糖基,底物11、12、13、14则分别在C20的6'-OH位点多一个葡萄糖、阿拉伯糖、木糖和芹糖基。UGT91Hs对具有额外糖基的底物表现出更高的活性,这暗示额外的糖基可能增强了底物与酶之间的相互作用,促进了反应。(iii)底物的不同立体构型也显著影响UGT91Hs的活性。底物8和9在C20位分别具有右旋(20R-)和左旋(20S-)构型,而UGT91Hs对具有S构型的底物9表现出更高的活性。
此外,为了表征UGT91Hs对糖基供体的底物特异性,我们使用各种UDP-糖(包括UDP-Glc、UDP-Gal、UDP-Xyl和UDP-GlcA)测试了上述30种受体底物。只有部分UGT91Hs在使用UDP-Xyl作为糖基供体时对底物1-5和9-15显示出活性(图S39-S43,S45-S51)。其中,UGT91H4、UGT91H6和UGT91H8在使用UDP-Xyl时表现出较高的转化率(表S2)。特别是UGT91H4和UGT91H8更倾向于对还原端为GlcA的二糖链底物进行糖基化,1-5的转化率超过20%。UGT91H6则倾向于对还原端为Glc的二糖链底物进行糖基化,9-15的转化率超过75%。然而,使用UDP-Glc、UDP-Gal和UDP-GlcA时未观察到活性。鉴于已报道的UGT91H4、UGT91H9和UGT91H11对糖基供体的底物特异性【12】,UGT91Hs对糖基供体表现出严格的特异性,最倾向于使用UDP-Rha,但只有一部分UGT91Hs能够利用UDP-Xyl和UDP-Glc。
UGT91H酶的祖先酶重建

如前所述,鉴定出的UGT91H酶显示出相当严格的底物特异性,这限制了它们在多种糖类广泛底物糖基化中的应用。我们决定利用祖先序列重建(ASR)获取UGT91H亚家族的祖先酶,期望其具有更广泛的底物范围。最终,我们从UGT91H亚家族的系统发育树中推断出7个不同的祖先节点(图3a,表S3)。首先选择了最古老的酶A1(即UGT91H_A1),它全面涵盖了UGT91H亚家族序列的特征。
我们还选择了UGT91H_A2、UGT91H_A3和UGT91H_A4,这些酶在系统发育树上的位置位于UGT91H_A1和UGT91H8之间(与UGT91H_A1的序列同一性为84.23%)。随后,将这四种祖先酶在大肠杆菌中表达并纯化,并使用UDP-鼠李糖作为糖基供体,测试它们对底物1、2和17的活性。UGT91H_A1、UGT91H_A2和UGT91H_A3保留了类似现存UGT91H酶催化底物1和2糖基化的能力(图3b和S52)。此外,它们还表现出对底物17的糖基化能力,而现存的UGT91H酶无法催化底物17。特别是,UGT91H_A1对底物17的转化率高达98.7±0.2%,表明位于UGT91H亚家族系统发育树根部的最古老的UGT91H_A1具有扩展现存UGT91H酶底物谱的潜力。我们测试了UGT91H_A1对底物1-30的底物范围,使用UDP-鼠李糖作为糖基供体。UGT91H_A1仅对在C3-OH处具有二糖链的三萜化合物表现出活性(图S2,S11和S19-S30)。与现存的UGT91H酶相比,UGT91H_A1对底物2、7、9、10、12、13、14和15显示出更高的转化率。令人惊讶的是,UGT91H_A1对底物16、17和18表现出超过80%的转化率,这些底物的二糖链C3-OH上带有阿拉伯糖(Ara)作为第一个糖基,而现存的UGT91H酶无法识别这些底物(图S53-S55)。此外,UGT91H_A1在糖基供体方面表现出更广泛的底物范围,如表S4所示。具体来说,UGT91H_A1不仅能像现存UGT91H酶一样催化底物5、9-15的木糖基化,还能催化底物7、16-18(图S43-S51,S56-S58)。此外,当UDP-葡萄糖(UDP-Glc)作为糖基供体时,UGT91H_A1能够催化底物16-18的葡糖基化反应,产物的[M-H]^-离子比底物高162 Da(图S59-S61)。UGT91H_A1表现出的这种罕见的葡糖基化活性可能是由于其序列特征与UGT91H9相似,而UGT91H9是唯一已知能够催化底物C2"-OH处葡糖基化的UGT91H酶,这表明ASR可以作为开发高效UGT酶的有效工具。为了验证UGT91H_A1产生的糖苷结构,我们选择底物17作为代表底物,通过NMR分析确定产物17a、17b和17c中糖基的连接位置(图S62-S82)。C2"-OH处引入的糖基通过C-2"和H-1'''的HMBC交叉峰得到确认,这表明UGT91H_A1很好地保持了对C2"-OH的严格区域选择性。我们的发现进一步支持了我们早先的推断,即UGT91H酶演化为负责在C2"-OH处添加一个糖基,用于合成带有线性三糖的三萜糖苷。此外,我们测定了UGT91H_A1对底物17使用各种UDP-糖的动力学参数(表2和图S83)。UGT91H_A1在使用UDP-鼠李糖时的催化效率(0.123 μM^-1▪s^-1)高于使用UDP-木糖(0.053 μM^-1▪s^-1)和UDP-葡糖(0.048 μM^-1▪s^-1)。因此,祖先酶UGT91H_A1不仅表现出普遍较高的活性,还扩展了糖受体和供体的底物范围,使其成为合成带有线性三糖的三萜糖苷的理想生物催化剂。
利用祖先酶扩展现存UGT91H酶的底物范围
根据经验,N端附近的组氨酸是大多数植物来源UGTs的催化残基。【17】通过与OsUGT91C1(PDB: 7ES1,唯一报道晶体结构的UGT91家族酶)进行序列比对,我们推测H29是UGT91H_A1的催化残基,D130作为辅助残基,对应于OsUGT91C1的H27和D128(图S84)。【18】随后,将H29和D130突变为丙氨酸后,UGT91H_A1的活性完全丧失,证实了保守的催化二联体(His-Asp)对于UGT91H_A1功能至关重要(图S85)。此外,正如表S5所示,UGT91H_A1催化产生的糖基化产物17a、17b和17c中的新形成的糖苷键相对于UDP-糖中的糖苷键均发生了反转。这一发现表明UGT91H_A1采用了SN2亲核取代反应机制,【19】其中H29和D130形成催化二联体,去质子化底物17的C2"-OH,生成SN2攻击UDP-Rha的亲核体(图4a)。
祖先酶UGT91H_A1与现存的UGT91H酶具有较高的序列相似性(66.03%-84.23%)(表S6)。与UGT91H8的序列同一性最高,达到84.23%。然而,UGT91H_A1相比UGT91H8展现出更广泛的底物谱,尤其是对底物17的催化能力。为识别影响UGT91H_A1底物特异性的关键残基,我们首先通过AlphaFold2对UGT91H_A1的结构进行建模。模型质量评估结果表明,UGT91H_A1的模型结构适合进行后续分析(图S86)。UGT91H_A1与底物17和UDP-Rha的分子对接显示,H29、N157、S298、E299、A354、H371和Q396与底物17和UDP-Rha形成了氢键(图S87)。这些残基在UGT91H8中也是保守的(图S88)。因此,推测在UGT91H_A1中可能存在一些非保守的关键残基影响其底物特异性。结合非保守残基与底物17和UDP-Rha之间的距离,我们推测F49、K100/I101/Y102、A133/D134、I158、L202/E203、R212/T213/A214/S215和M380可能是影响底物特异性的关键氨基酸(图4b)。
尽管UGT91H8与UGT91H_A1的序列同一性为84.23%,但UGT91H8缺乏催化底物17糖基化的能力,而UGT91H_A1对17的转化率达到了98.7±0.2%。因此,我们将UGT91H_A1中上述七个非保守残基与UGT91H8的对应区域进行了互换,旨在赋予UGT91H8催化17糖基化的能力。成功在大肠杆菌中表达并纯化了七个UGT91H8突变体,分别命名为UGT91H8_1mu(L50F)、UGT91H8_2mu(M99K/N100I/R101Y)、UGT91H8_3mu(S132A/G133D)、UGT91H8_4mu(K157I)、UGT91H8_5mu(F200L/T201E)、UGT91H8_6mu(G209R/M210T/A211A/G212S)和UGT91H8_7mu(K377M)(图S1)。只有突变体UGT91H8_6mu表现出新的活性,能够催化17的鼠李糖基化,转化率为15.9±0.5%,并且对17的木糖基化也达到了7.7±0.8%的转化率(图4c和图S89)。此外,底物范围测试表明,突变体UGT91H8_6mu对底物16和18也表现出新的活性,这些底物的分子结构与17相似,C3-OH处带有二糖,第一个糖基为阿拉伯糖(Ara)。LC-MS/MS分析验证了UGT91H8_6mu对16、17和18的新活性(图S90-S91和表S7)。该突变体中的四个突变残基形成了一个环状结构(命名为RTAS环)。为了研究该环对UGT91H亚家族中底物特异性的普遍影响,RTAS环被分别替换为其他UGT91Hs中的对应区域。成功生成了五个突变体,包括UGT91H5M、UGT91H6M、UGT91H7M、UGT91H12M和UGT91H13M。其中,突变体UGT91H7M获得了催化17鼠李糖基化的新活性,转化率为12.6±0.3%,而其他突变体对17仍无活性(图S92-S93)。最后,将UGT91H_A1的RTAS环替换为UGT91H8的对应区域(G209/M210/A211/G212,即GMAG环),生成了突变体UGT91H_A1_6mu(R212G/T213M/A214A/S215G)。与UGT91H_A1相比,突变体UGT91H_A1_6mu对16和17的活性显著降低。具体而言,UGT91H_A1_6mu对16和17的鼠李糖基化转化率分别从81.8±3.4%降至33.8±3.2%和从98.7±0.2%降至62.4±5.4%(图S94)。这些发现表明,RTAS环在调节UGT91H_A1的底物特异性中起重要作用,同时也是工程化UGT91Hs底物特异性的一个关键区域。
研究RTAS环对底物特异性的影响机制

为了阐明RTAS环在反应中的作用,我们使用分子动力学(MD)模拟和量子力学(QM)计算比较了UGT91H8_6mu和UGT91H8催化的糖基化反应的反应性。通过AlphaFold2生成UGT91H8_6mu和UGT91H8的模型结构,模型质量评估结果适合进一步分析(图S95-S96)。根据UGT91H_A1和UGT91H8的序列比对,确定H30和D129是UGT91H8及其突变体UGT91H8_6mu的催化二联体(图S84)。然后,将UDP-Rha和底物17分别对接到UGT91H8_6mu和UGT91H8的结构中,并使用适当的酶-底物复合物构象进行后续的MD模拟。根据蛋白质骨架和配体的均方根偏差(RMSD)值,复合物结构在70 ns MD模拟后达到稳定状态(图S97)。在UGTs的催化过程中,糖受体的氧原子发生去质子化,并与UDP-糖的变亚甲基碳原子形成新的糖苷键。因此,必须保持两个底物之间的稳定近距离。
在UGT91H8_6mu复合物的MD模拟过程中,底物17的C2"-OH的氧原子与UDP-Rha的变亚甲基碳原子之间的距离(O-C距离)在约60 ns的MD模拟后缩短并稳定在小于6 Å的范围内(图5a)。相反,在UGT91H8复合物的MD模拟中,O-C距离在约85 ns的MD模拟后逐渐增加到超过8 Å。结果表明,UGT91H_A1可能创造了一个相对稳定的环境,以保持17和UDP-Rha之间的更近距离,从而促进17的鼠李糖基化反应,而UGT91H8未能实现这一点。此外,选取了150 ns MD模拟的代表性构象,分析UGT91H8或UGT91H8_6mu与17和UDP-Rha的相互作用(图S98-S99)。在UGT91H8复合物中,七个残基(T91、N100、S295、E296、H368、D392和Q393)与底物形成了氢键,而四个氨基酸(F160、L203、F213和Q353)参与了与底物的疏水相互作用。在UGT91H8_6mu复合物中,九个残基(H30、N100、S212、E296、H368、S371、E376、D392和Q393)与底物形成了氢键,而六个残基(Y32、I151、F130、F160、L203和L215)参与了与底物的疏水相互作用。因此,UGT91H8_6mu与底物形成了更多的相互作用,促进了底物识别和催化。
值得注意的是,在UGT91H8_6mu复合物的MD模拟过程中,RTAS环中的S212残基逐渐接近17上二糖链的鼠李糖部分,环发生摆动后,S212羟基的氢原子与17中鼠李糖基的氧原子形成了一个氢键(O-H...O),在约75 ns的MD模拟后稳定在2.0-3.5 Å范围内(图5b)。相比之下,在UGT91H8复合物的MD模拟过程中,GAMG环的对应残基未能与17形成氢键,G212残基氨基中的氢原子与17上鼠李糖基的氧原子之间的距离(NH-O)在整个MD模拟过程中保持在8 Å以上。
此外,我们通过QM计算比较了UGT91H8_6mu和UGT91H8在不同反应状态下的相对能量(图5c)。从MD模拟中选择UGT91H8_6mu和UGT91H8的150 ns瞬时构象作为初始状态(酶与UDP-Rha和17结合),通过对接得到最终状态(酶与UDP和鼠李糖基化的17结合)。首先,我们计算了包含17、UDP-Rha、H30、D129和R209/G209、T210/M210、A211、S212/G212的UGT91H8_6mu和UGT91H8的小模型(图S100-S101)。与实验结果一致,UGT91H8_6mu的小模型中,17的鼠李糖基化反应的初始状态(-94.5 kcal/mol)和最终状态(-139.0 kcal/mol)的相对能量低于UGT91H8的对应状态(表S8)。为了提高计算精度,并避免小模型计算结果的偶然性,我们还将N100、E296(参与氢键作用)和F160、L203(参与疏水相互作用)加入到大模型中(图S102-S103)。UGT91H8_6mu的初始状态(-127.2 kcal/mol)和最终状态(-175.2 kcal/mol)的相对能量显著低于UGT91H8的对应状态(表S9)。这些结果表明,由于较低的结合能,UGT91H8_6mu更倾向于催化17的鼠李糖基化反应。
UGT91H_A1结合UDP-Rha再生系统合成带有线性三糖的三萜糖苷2a和17a

UDP-糖的高成本和副产物UDP的抑制作用显著限制了大规模糖苷生产的进展。【20】令人鼓舞的是,将蔗糖合酶(SUS)、UDP-鼠李糖合酶(RHM)和双功能鼠李糖合酶(NRS/ER)结合,构建自给式UDP-Rha再生系统,已被报道作为一种策略,可从价格低廉的蔗糖中提供用于鼠李糖苷合成的UDP-Rha。【21】值得注意的是,在RHM的催化过程中,UDP-葡萄糖经过其N端区域催化的4,6-脱水作用,导致NAD+被还原为NADH。随后,UDP-4-酮-鼠李糖在其C端区域催化下发生还原反应,导致NADPH被氧化为NADP+(图S104)。【22】NRS/ER能够催化NADH的氧化以回收辅因子NAD+。【21a, 23】基于我们之前的研究,【24】我们将GuSUS1-Δ9、AtRHM2、AtNRS/ER和UGT91H_A1结合,构建了一个四酶UDP循环系统,用于催化2和17的鼠李糖基化(图S105)。在此系统中,GuSUS1-Δ9、蔗糖、UDP和PBS(pH 5.5)最初在50℃下孵育1小时。随后将反应体系的pH调整为7.5,加入AtRHM2、AtNRS/ER、UGT91H_A1、NAD+和底物2或17,并在35℃下孵育12小时。经过12小时反应后,2a的产量达到160.1±0.6 μM,2的转化率增加到80.1±0.3%。同样,17a的产量在12小时反应后达到122.7±8.3 μM,转化率为61.3±4.1%(图6)。尽管在UDP-Rha再生系统中生成了中间产物UDP-葡萄糖,且UGT91H_A1具备使用UDP-葡萄糖对17进行糖基化的能力,但在反应系统中未检测到葡萄糖基化产物17c(图S106)。总的来说,结合祖先酶UGT91H_A1的UDP-Rha再生系统在高效生产带有线性三糖的三萜糖苷方面具有重要潜力。
通过基于系统发育的UGT酶挖掘策略扩展UGT91H亚家族

如前所述,已报道的UGT91H酶数量非常有限,极大限制了多样化的带有线性三糖的三萜糖苷的合成。本研究表明,UGT91H酶倾向于具有相似的功能。因此,使用已表征的UGT91H作为模板,系统发育分析可能是从庞大的植物基因组数据中高效定位更多UGT91H成员的可行方法。基于这一思路,我们利用9种已知的UGT91H酶的序列作为模板,从8种富含三萜糖苷的豆科植物中发现了共23个候选UGT(表S10)。其中,19个UGT属于UGT91H亚家族,4个UGT属于一个新的UGT91BQ亚家族。这表明基于系统发育的UGT挖掘策略在快速扩展UGT91H亚家族方面是有效的。系统发育分析表明,来自红三叶草(Trifolium pratense)的PNY09112.1(UGT91H19)和PNX78912.1(UGT91H21),以及来自地下三叶草(Trifolium subterraneum)的GAU40116.1(UGT91H20)可能具有针对三萜化合物C2"-OH位点的糖基化功能,因为它们与已表征的UGT91H酶在同一进化分支上(图7a)。
随后我们表征了UGT91H19、UGT91H20和UGT91H21的活性,并探索了它们在扩展UGT91H酶底物范围方面的潜力。UGT91H19、UGT91H20和UGT91H21对底物1和2表现出活性,但对底物16-17和19-22没有活性(图7b,图S107-S112和表S11)。值得注意的是,UGT91H19和UGT91H20不仅催化了底物1的鼠李糖基化、木糖基化和葡萄糖基化,还表现出催化1的半乳糖基化的活性,生成了产物1a、1b、1c和1d。尽管1的半乳糖基化转化率只有6.8 ± 0.1%,这种活性是迄今为止报道的UGT91H酶所不具备的。未来,可以探索对UGT91H19和UGT91H20进行分子改造,以增强其对底物的半乳糖基化能力,从而实现多种类型的三萜糖苷的高效合成。因此,通过蛋白质序列比对和系统发育分析从植物基因组中高效挖掘UGT91H酶的策略,可以进一步扩展到其他UGT亚家族的探索。
结论
总而言之,UGT91H酶被证明是三萜类化合物2"-O-糖基化的重要生物催化剂。在此基础上,我们应用祖先序列重建(ASR)来拓宽UGT91H亚家族的底物范围。祖先酶UGT91H_A1展示了催化底物16、17和18的C2"-OH糖基化的新活性。通过使用现存的UGT91H酶和祖先酶UGT91H_A1,我们首次酶促合成了23种带有线性三糖的三萜糖苷,包括鼠李糖基化衍生物1a、2a、4a、7a、15a、16a、17a和18a,木糖基化衍生物1b、2b、4b、5b、7b、9b、10b、11b、12b、13b、14b和15b,以及葡萄糖基化衍生物16c、17c和18c。我们发现UGT91H_A1中的RTAS环影响底物特异性,将此RTAS环转移到UGT91Hs的相应位置产生了UGT91H8_6mu和UGT91H7M突变体,它们对新底物17表现出新的特异性。MD和QM计算表明,RTAS环能够影响酶与底物之间的相互作用和结合能,从而调节底物特异性。此外,结合逐步的UDP-Rha再生系统,祖先酶UGT91H_A1能够经济高效地合成鼠李糖基化三萜类化合物2a和17a。最后,我们开发了一种基于系统发育的UGT挖掘策略,成功从8种豆科植物中鉴定出UGT91H家族的19个新成员。这一策略可以轻松应用于其他UGT的快速挖掘。本研究为三萜类化合物的C2"-OH糖基化提供了高效的生物催化剂,并强调了ASR在拓宽现存UGT91Hs底物特异性方面的价值,促进了更多带有线性三糖的三萜糖苷的合成。

![[AutoSar]BSW_Diagnostic_005 RoutineControl service (0x31)介绍](https://i-blog.csdnimg.cn/direct/3e846ae973f64a61844f4ce815e86fa0.png)