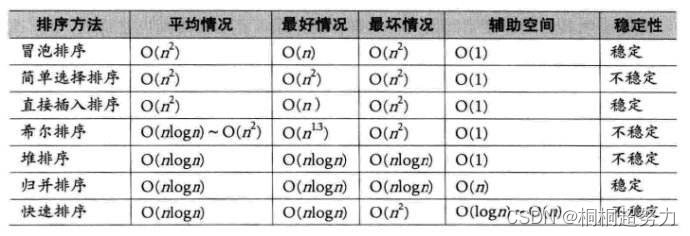
非人灵长类动物(NHP)是人类的近亲,为宿主-微生物互作的研究提供了一个很好的例子。近年来研究主要集中在野生灵长类动物的肠道微生物群,这将有助于了解灵长类及其肠道微生物群的进化,但仍然缺乏关于野生种群肠道微生物群的基本信息。
今日分享一篇发表在《Computational and Structural Biotechnology Journal》期刊上(IF=6.155)的论文,研究发现野生觅食和食物供应的云南金丝猴种群的肠道微生物群存在显著差异,野生种群的长期食物供应导致其肠道微生物组成、功能甚至抗生素耐药性的动态变化。
实验设计
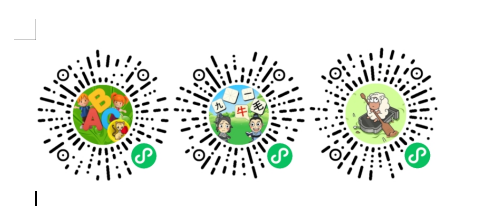
利用16S rRNA技术、宏基因组和宏基因组binning探究比较了中国云南省威西县云南金丝猴种群的肠道微生物组成和功能,这些种群的饮食完全基于野生觅食或定期补充人类提供的食物。

图1云南金丝猴的研究区域和饮食
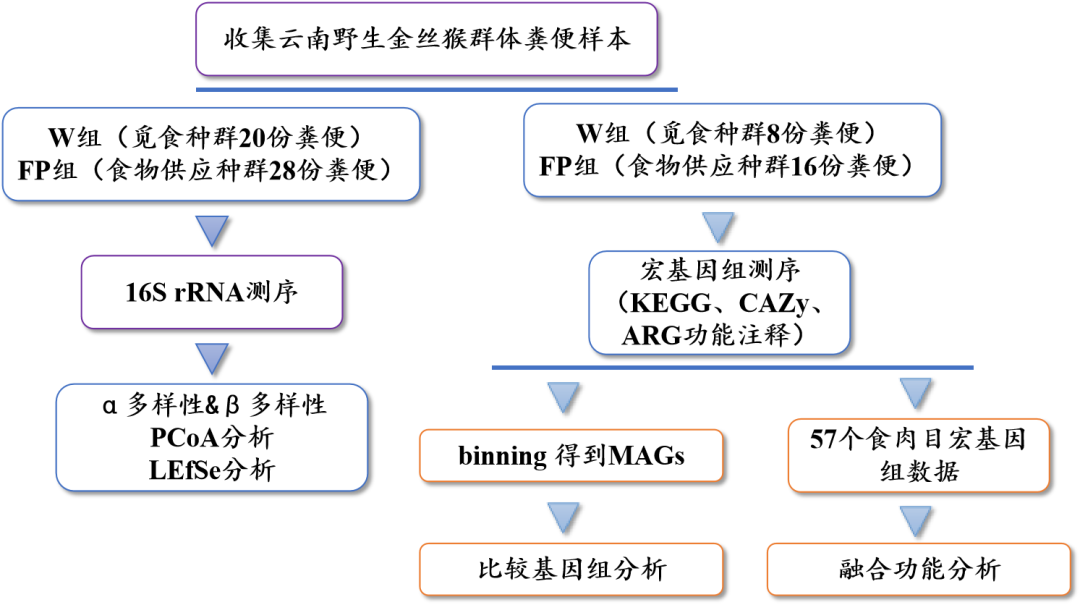
图 研究设计
主要研究结果
1、野生觅食和食物供应种群肠道微生物组成的显著差异
使用16S rRNA技术,发现野生觅食(W组)和食物供应(FP组)云南金丝猴种群的肠道微生物群落存在显著差异。变形菌门、拟杆菌门和螺旋菌门的相对丰度在W组中显著高于FP组(图2A和2B)。FP组中厚壁菌门、放线菌门和纤维杆菌门的相对丰度显著高于W组(图2A和2B)。α多样性中,食物供应种群的香农指数显著高于野生觅食种群。更多样化的食物供应可能会导致更高度多样化的肠道微生物群(表1)。PCoA分析证明了W和FP组之间肠道微生物群落的显著差异(图2C)。此外,与FP组相比,W组的微生物群落组成更相似。
这项研究中,在同一自然区域内的野生觅食和食物供应种群之间,肠道微生物群组成存在差异。这种差异可能与饮食组成的差异有关,例如,竹笋摄入比例在W群体中较高,但在FP群体中较低(表1)。大多数竹笋含有高比例的氰化物。食竹肠道微生物群显示出高比例的变形菌门菌群(如假单胞菌),这与竹子中氰化物的解毒有关。厚壁菌门和纤维杆菌门菌群的显著富集可能参与FP中高碳水化合物食物的消化有关。在W组观察到食虫行为,由于昆虫的甲壳素含量很高,猜测与FP群体相比,编码参与几丁质降解的酶的基因将在W群体中显著富集。
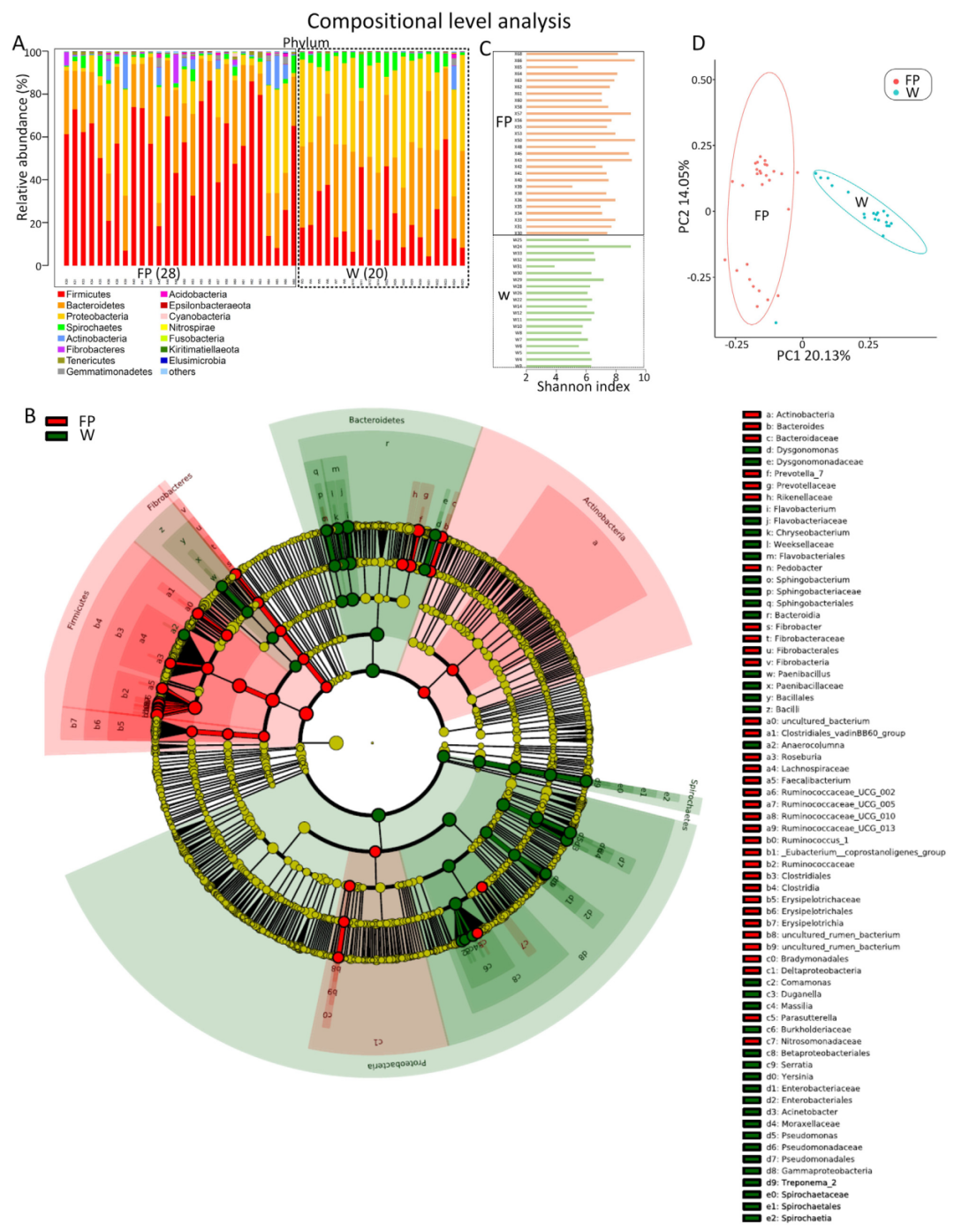
图2 W和FP组中的肠道微生物群落
表1 W组和FP组的食物贡献率
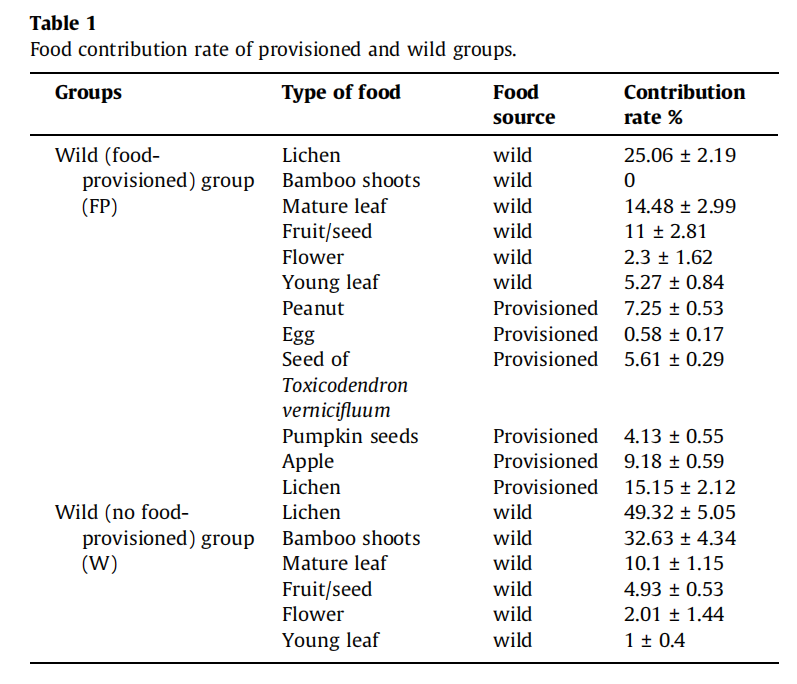
2、W和FP组之间肠道微生物群的功能差异
分析了24个粪便宏基因组,发现W组中来自变形菌门和拟杆菌门的属居多,而来自厚壁菌门、拟杆菌门和纤维杆菌门的属在FP组中富集(图3A)。PCA分析表明,觅食和食物供应群体之间的肠道微生物功能存在显著的功能差异(图3B)。使用KEGG途径丰度的Lefse分析表明,许多碳水化合物代谢和能量代谢途径的丰度在FP组的肠道微生物群中富集(图3C)。
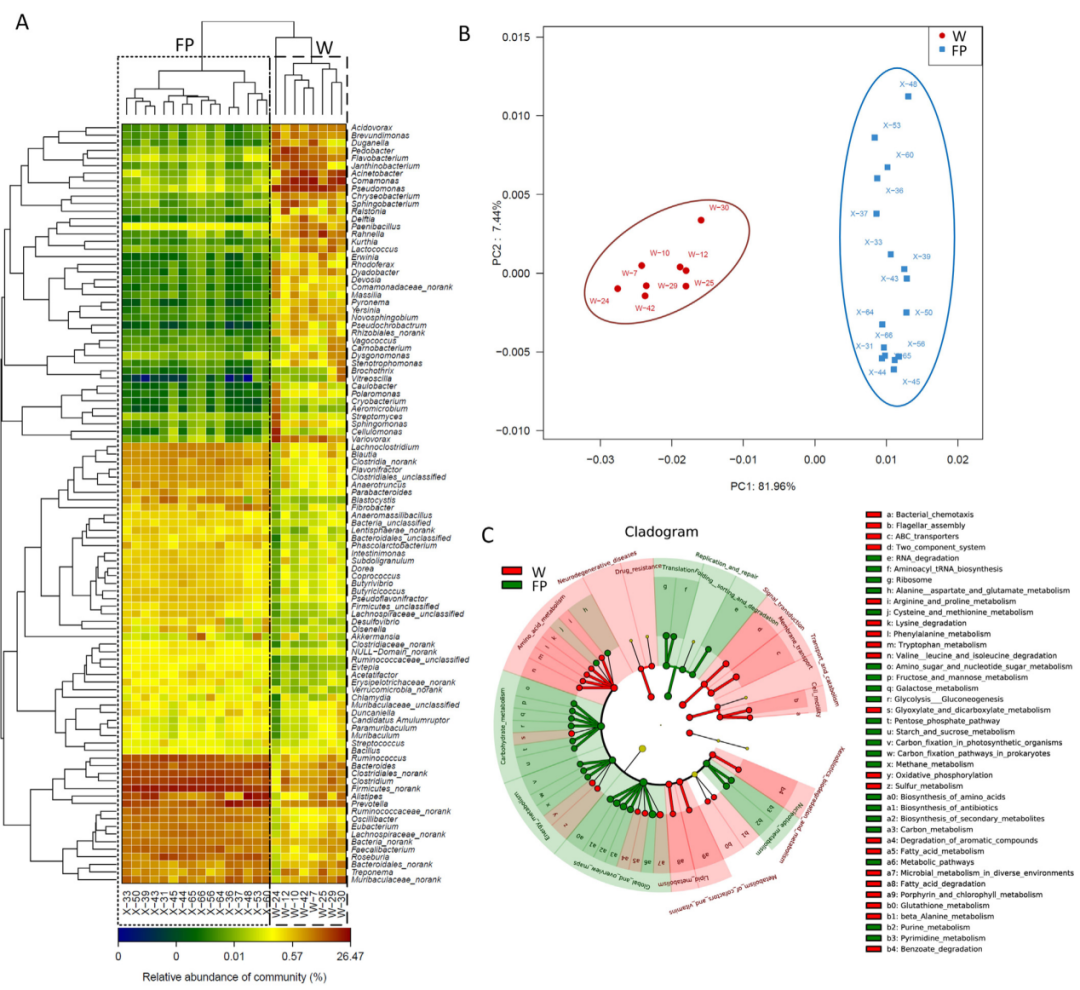
图3 24个宏基因组的功能分析
FP组中参与碳水化合物代谢的六条KEGG功能通路的主要肠道微生物属包括普氏杆菌、瘤胃球菌属、Alistipes、梭杆菌属等(图4)。肠道微生物功能的差异表明肠道微生物组对这些人群之间饮食差异的反应。
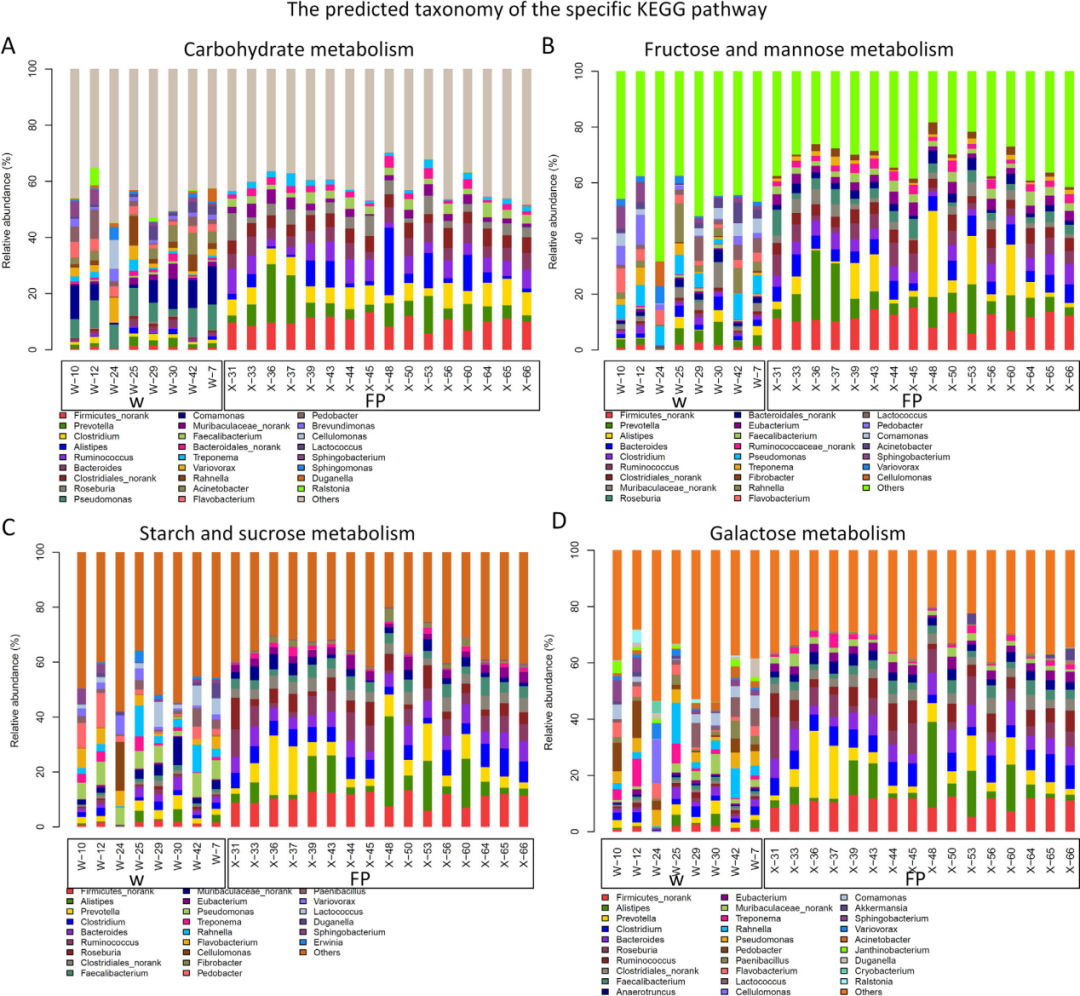
图4 24个宏基因组预测的KEGG功能分类
CAZy分析进一步证实了上述发现(图5)。例如,22个GH家族(糖苷水解酶)的丰度在FP组中显著高于W(图5A)。其中GH2和GH43家族推测来自FP中的普氏杆菌、拟杆菌门、瘤胃球菌、梭杆菌等(图5B)。GH23(包括G型溶菌酶和几丁质酶)是W中丰度最高的GH家族,显著高于FP群体(图5A)。W群体中编码假定几丁质酶的基因丰度显著高于FP群体(图6A)。
随后比较了编码主要参与竹子降解的特定酶的基因。编码参与该物质降解的酶的基因(包括硝化酶(TST)、硫代硫酸盐硫转移酶(glpE)等),与FP群体相比,W群体的肠道微生物群显著富集(图6B),编码基因主要来源于变形菌门,如W群体中的Comamonas、假单胞菌、不动杆菌、Rahnella和Variovorax(图6B)。因此,这也可能反映了云南金丝猴肠道微生物的可塑性。
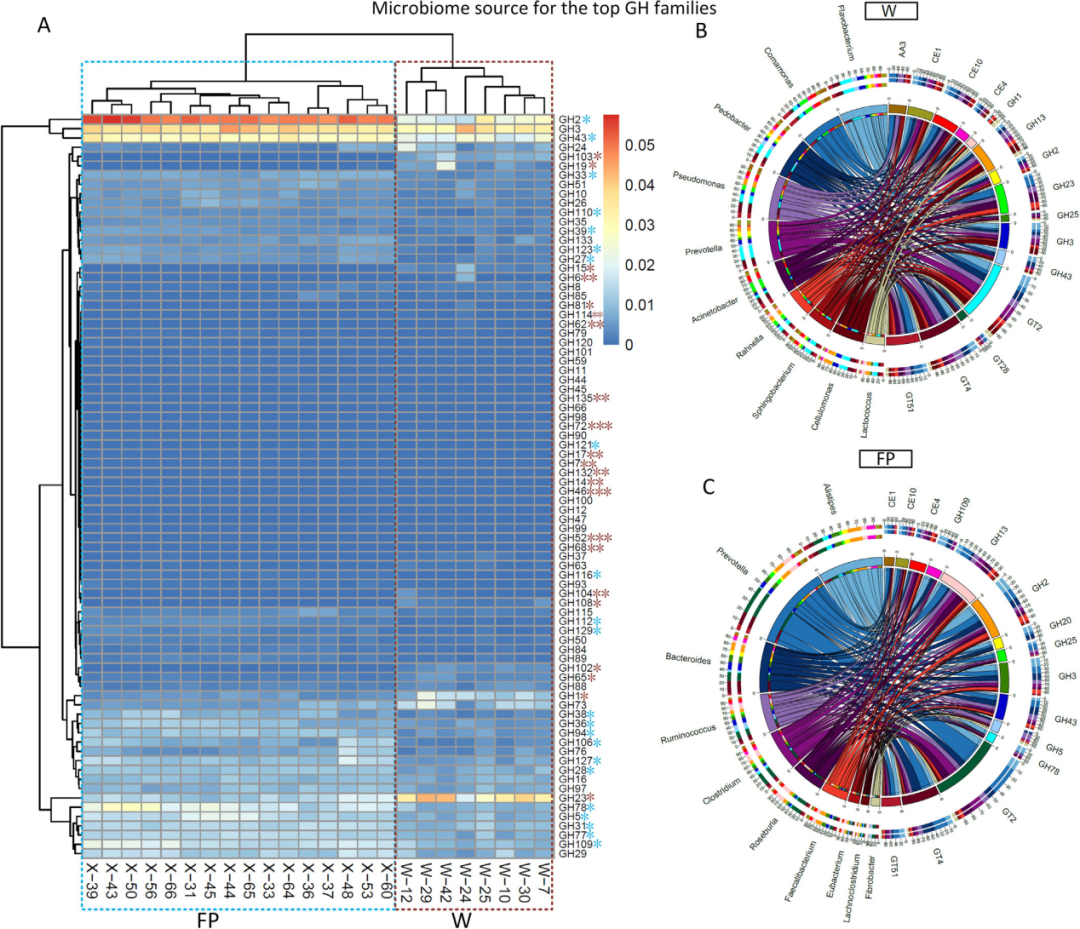
图5 24个宏基因组的CAZy分析
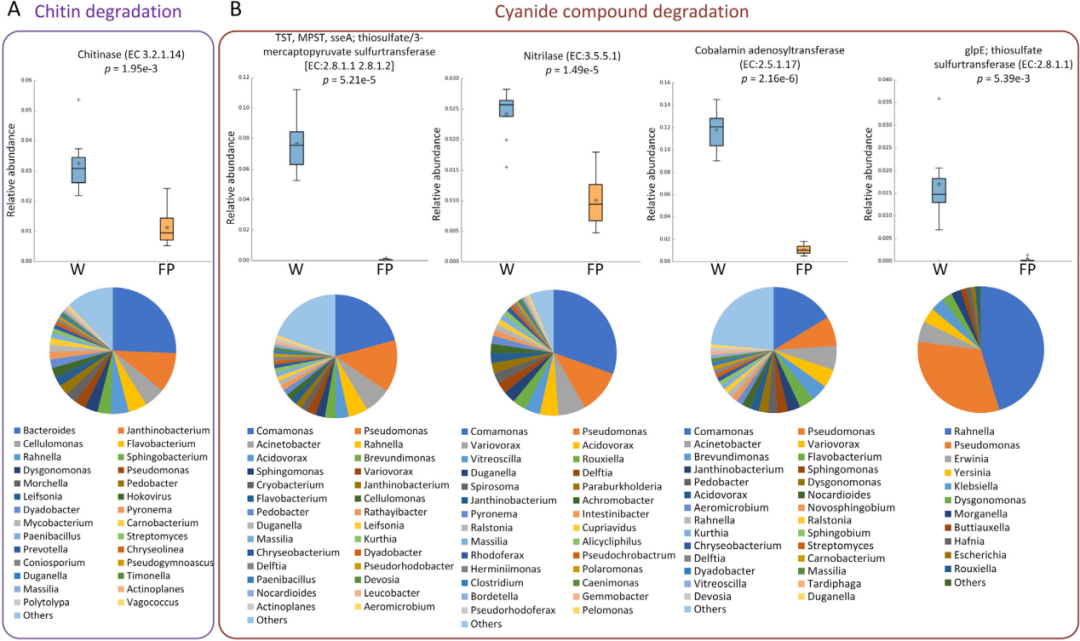
图6 编码参与几丁质和氰化物化合物降解的特定酶的基因
在W和FP云南金丝猴种群的肠道微生物群发生深刻变化后,作者探索对宿主健康的潜在影响。通过宏基因组对抗生素抗性基因(ARG)谱进行了预测,发现这两组之间存在显著差异(图7A和7B)。W组的肠道微生物群富含多种耐药亚型(例如,mul_acrB、mul_ mdfA等),这些亚型可能来源于假单胞菌属、不动杆菌属、Rahnella、Comamonas、Variovorax和Janthinobacterium(图7C)。FP中的肠道微生物群富含四环素亚型(如tet_tetQ、tet_ ykkD)和万古霉素抗性(如van_vanU、van_ vanY等),这些亚型可能源自拟杆菌属、普氏杆菌属和瘤胃球菌属(图7D)。
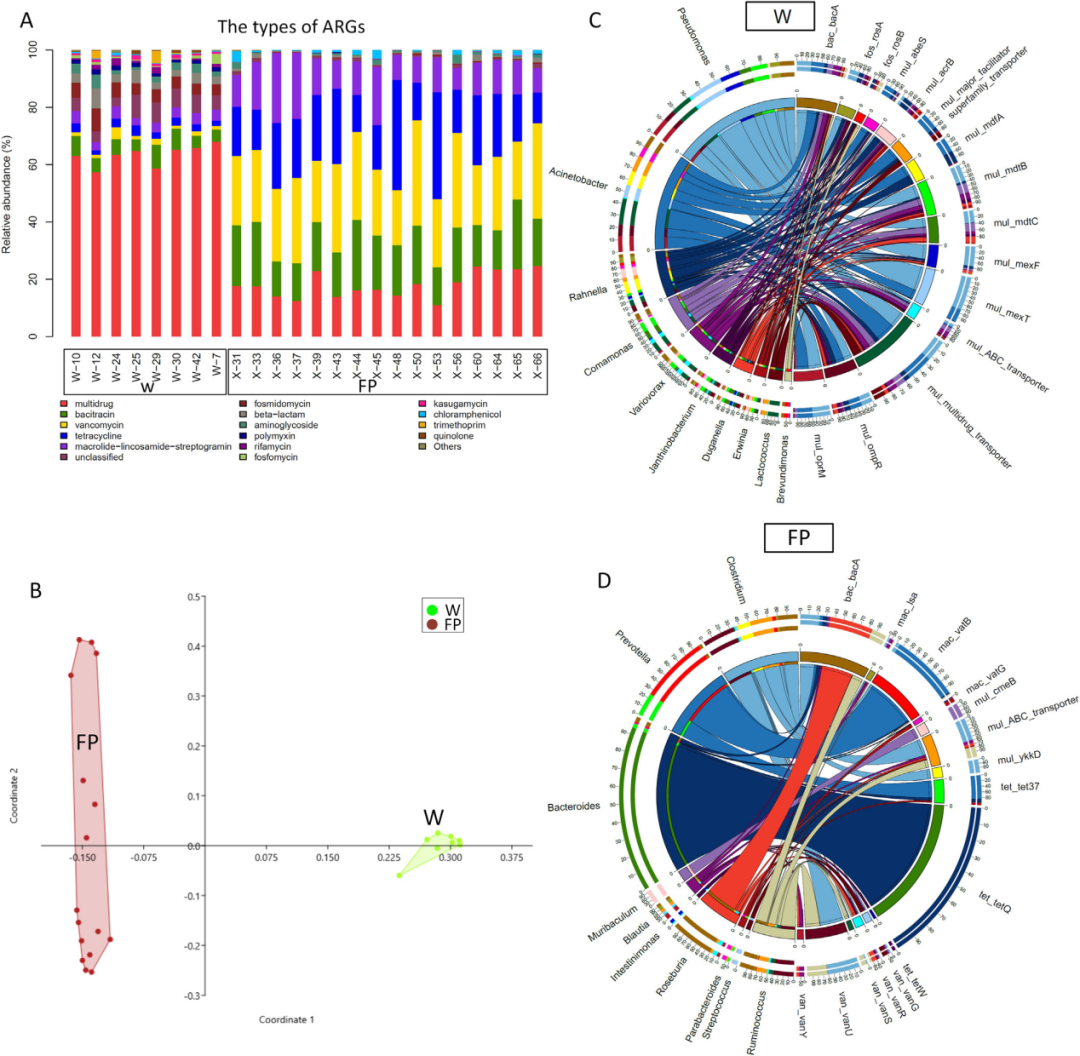
图7 W和FP组的24个宏基因组中的抗生素抗性基因(ARGs)谱
3、MAG结果证实富集的基因与降解特定饮食高度相关
宏基因组binning得到了88个高质量MAGs,比较基因组分析表明,这些MAGs的相对丰度在W组和FP组之间不同(图8)。来自变形菌门、放线菌门和拟杆菌门的MAGs的相对丰度在W组中较高,但在FP中罕见。MAGs中编码硝化酶(TST)、硫代硫酸盐硫转移酶(glpE)等的基因主要分布在变形菌门和拟杆菌门。编码假定几丁质酶的基因主要分布在W组中,证明了W和FP组之间的MAG相对丰度存在差异,这可能与不同的饮食有关。
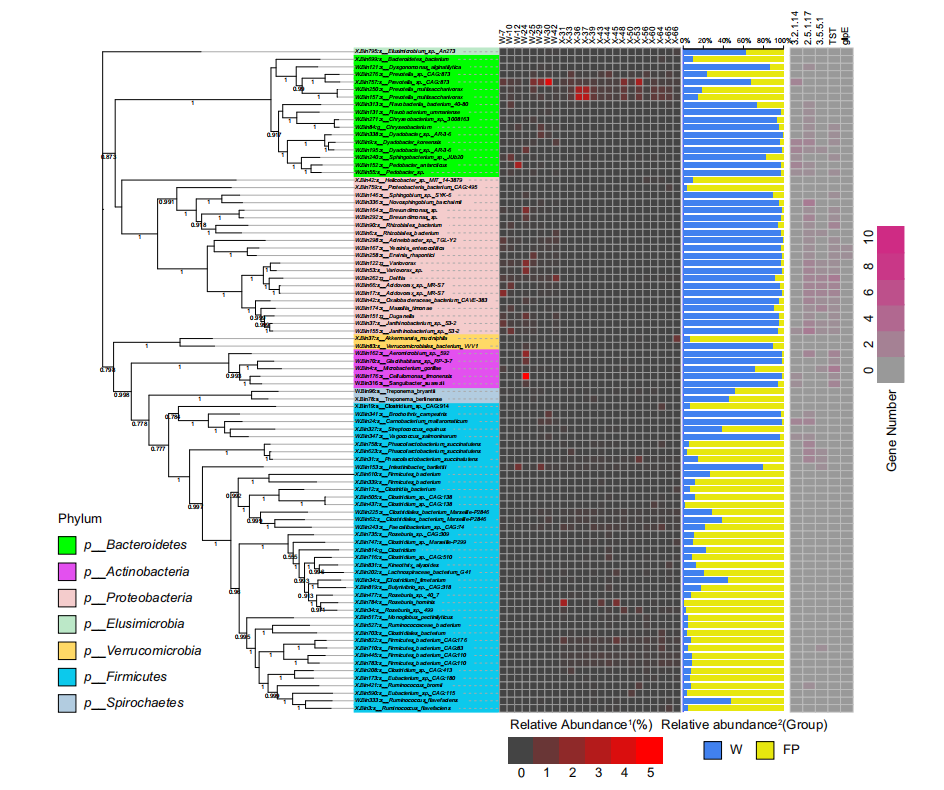
图8 对MAGs的系统发育分析
4、云南金丝猴与食竹熊猫肠道微生物群的功能融合
野生觅食云南金丝猴肠道微生物功能的一个主要特征是编码参与竹子中氰化物降解酶的基因丰富。基于之前发表的57个宏基因组数据,包括9个CA(食肉肉食动物)、12个HE(食草动物)、10个OC(杂食肉食动物)和10个GP(大熊猫)和6个RP(小熊猫),推测野生觅食(竹笋摄入量高,不提供补充食物)金丝猴种群与野生食竹大熊猫的肠道微生物群具有功能趋同性。PCoA分析和分级聚类证明,两种动物肠道微生物功能高度相似(图9A和9B),证明了尽管它们属于不同的哺乳动物目(灵长目与食肉目),但饮食在塑造肠道微生物群中起着重要作用,饮食驱动不同哺乳动物肠道微生物群的趋同。

图9 云南金丝猴和食竹熊猫肠道微生物群使用81个宏基因组的功能融合分析
研究结论
本研究发现了野生觅食和食物供应的云南金丝猴种群的肠道微生物群存在显著差异,这可能与不同的饮食组成有关。野生觅食(未提供补充食物)种群与野生食竹大熊猫在肠道微生物群中功能趋同:例如与氰化物解毒有关的微生物和基因。长期给野生NHP种群供应食物导致肠道微生物组成、功能甚至抗生素耐药性的动态变化。野生NHP种群的抗生素耐药性概况将为将为其保护的提供最基本和重要信息。
参考文献
Functional convergence of Yunnan snub-nosed monkey and bambooeating panda gut microbiomes revealing the driving by dietary flexibility on mammal gut microbiome. Computational and Structural Biotechnology Journal, 2022.
往期推荐
《mSystems》比较宏基因组学探究海洋和陆地生态系统中磷酸盐分解代谢途径
极地土活性层和永久冻土的比较宏基因组学