《Protein Actions Principles and Modeling》-《蛋白质作用原理和建模》
本人能力有限,如果错误欢迎批评指正。
第四章:Protein Binding Leads to Biological Actions
(蛋白质的结合会产生生物作用)
-布朗棘轮在耦合结合的过程中产生定向运动
生物系统如何进行定向运动是一个有趣的研究问题。有些蛋白质是只向一个方向旋转的旋转马达。而有些蛋白质则像是沿着其他轨道形状蛋白往一个方向行走的线性马达。这些定向行动是为了将细胞内的分子运送到目的地,或以直接的方式将分子泵入细胞或使其离开细胞,或使肌肉进行收缩。因为细胞非常小,所以布朗运动会使这些细胞中的分子随意运动。那么生物分子是如何实现定向运动的呢?结合和解离与ATP到ADP的向下电流以及传导耦合,会导致定向运动的产生。因此,这种过程又称为布朗棘轮。

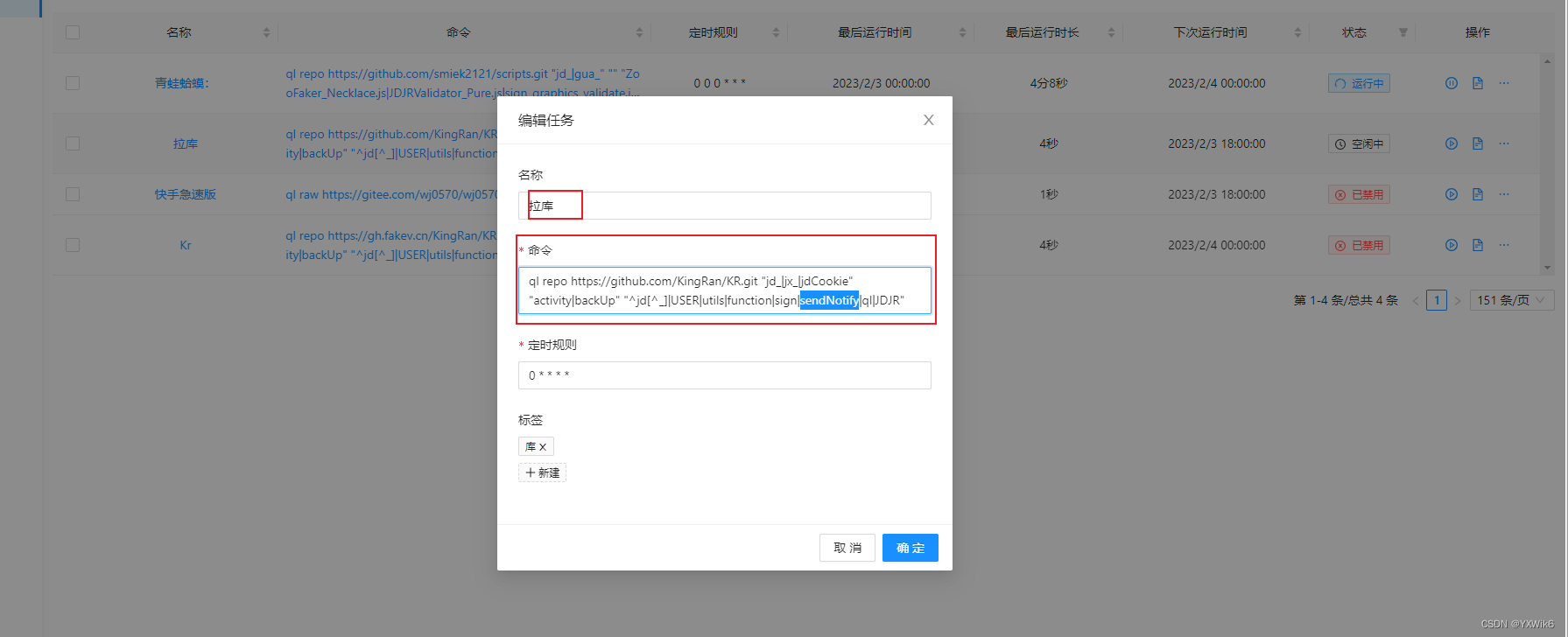
图4.18 在结合解离循环中布朗棘轮会导致定向运动。粒子以不对称的相互作用结合到轨道上,这里显示为“锯齿”作用。它脱离轨道时,粒子随机水平扩散。但是,当它重新结合到轨道上时,由于电势的不对称性,粒子更容易落入右边的能量最低点中,而不是左边的能量最低点中。高斯扩散曲线P (x)下的阴影表明,落入右最低点的概率高于落入左最低点的概率。
图4.18展示了布朗棘轮如何产生定向运动。让我们想象一下一个与具有锯齿形能量函数的轨道分子结合的粒子。最左边的图像(A)显示了在最小能量点上的结合粒子。(B)展示粒子从低能束缚态释放出来,该过程需要一些外部能量来源的输入,如ATP转化为ADP。(C)显示了未被束缚的粒子现在是如何随机地左右扩散的。(C)上的高斯分布展示了分子经过一段平均扩散时间的横向分布不存在偏向性的角度。(C)中的灰色阴影显示了这是如何发生的:因为锯齿能量函数是不对称的,粒子落入右边最低点的概率比落入左边最低点的概率要高。(D)显示,粒子通常落入到它开始处的右边的最低点中。因此,粒子与轨道的结合和分离的重复循环,加上每个循环的能量利用,导致粒子的平均向右运动。BOX4.6以解旋酶作为例子进行具体分析。解旋酶是一种沿着DNA单向滑动的马达蛋白,就像沿着轨道的火车一样,通过解开两条DNA链来分离它们。解旋酶可以实现DNA到RNA的转变和其他过程,如DNA修复和重组。
====================================================
BOX 4.6 解旋酶的定向运动
解旋酶是沿着DNA双螺旋结构(像轨道)移动的一种蛋白质,它可以将双链DNA解开成单链。在这里,我们只是考虑ATP的能量是如何被用来推动解旋酶沿着DNA的一个单一方向。图4.19显示了一个布朗棘轮样模型,其中解旋酶与DNA结合,同时伴有ATP水解为ADP,产生定向运动。
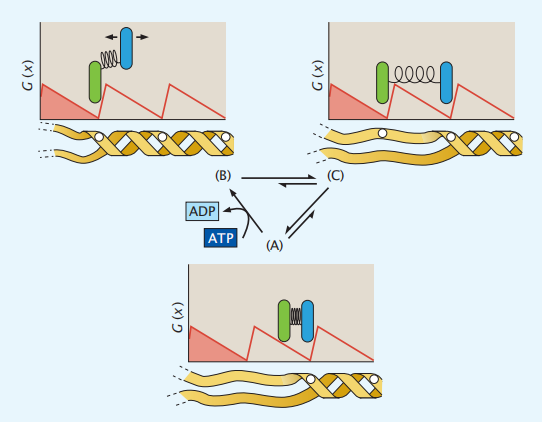
图 4.19 解旋酶的运动循环。解旋酶有两个亚基。(A)解旋酶与DNA结合。(B)一个亚基(蓝色)在ATP转化为ADP的驱动下解开结合,并向右移动一步,在此期间DNA被解开。C)另一个亚基(绿色)现在被第一个亚基拉到右边。整个解旋酶蛋白现在已经向右走了一步,然后重复该步骤。白点是标记解旋酶与DNA结合的前面和后面的位点。
====================================================
布朗棘轮运动的另一个例子是动力学校正(kinetic proofreading)。这是一个质量控制的分子过程,可以确保生物过程的准确性。如:RNA聚合酶将DNA转化为RNA,或者是将mRNA转化为蛋白质。在这样的聚合过程中,新的单体按照一个模板,被添加到一个不断生长的链上,形成正确的序列。在动力学校正中,通过聚合过程与结合耦合以及GTP-GDP的转变可以降低错误,如BOX4.7所示。
====================================================
BOX 4.7 动力学矫正可以降低蛋白质合成的复制错误
在蛋白质合成过程中,氨基酸被逐个地添加到一个不断增长的蛋白质链上。一种携带特定氨基酸的tRNA在细胞中扩散,并与引导蛋白质合成的核糖体上的信使RNA结合。这20个不同的tRNA分子试图与相同的核酸mRNA密码子(N)结合。然而最重要的是,只有正确的tRNA (C)最终才能结合其同源mRNA。在细胞中,这种选择具有高度准确的不正确的反密码子(I)与mRNA链结合的频率只有每10000个单位具有1个错误。然而,这些反密码子和密码子具有非常相似的结构和能量学。这种匹配是如何如此准确地进行的?这涉及两个作用:(1)正确的结合亲和度就占了大约100倍的选择性(图4.20),(2)激活的动力学过程,称为动力学校正,也占了100倍(图4.21)。
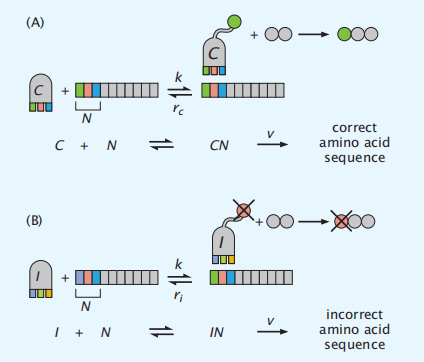
图4.20 一个mRNA密码子(N)与(A)的正确(或同源)tRNA(C)结合,或(B)与不正确的tRNA (I)结合。在mRNA结合tRNA(C或I)后,蛋白质链以v的速度拉长。实验表明,C的结合强度比i紧密100倍,但这并不足以进行高精度复制。动力学校正提高了准确性。
首先,考虑最简单的选择性机制:结合亲和度的差异。图4.20显示了一个快速的平衡结合步骤,然后是蛋白质链的以v为速度的延伸步骤所以,我们有 [CN] = KC[C][N]以及 [IN] = KI[I][N]。而正确和错误反密码子的结合亲和度KC和KI分别为

其中rC,ri分别是为正确,错误的tRNAs离开mRNA的解离速率。

图4.21 动力学校对将氨基酸添加到GTP结合. (A),密码子与反密码子正确匹配(B);密码子与反密码子错误匹配(B)。
这些反应具有大约相同的正向速率k,因为tRNAs是通过一种蛋白质(细菌中的EF-Tu)扩散到核糖体上的mRNA靶点的。这种扩散速率与tRNA没有太大的关系。相比之下,C和I的亲和度的不同是因为它们的解离率rC和ri不同。解离率受结合结构和能量影响。C与mRNA的结合更紧密,所以它脱落得更慢。选择性可由结合率的比率给出:

我们假设[C]≈[I]是因为2将0种不同的trna都将以大致相同的可得浓度进行计算。100的值来自于实验。公式4.42表明,正确的配体比不正确的配体的解离率小100倍。
现在,细胞过程通过一个额外的生化步骤实现了更高的选择性(见图4.21)。在细胞中,tRNA与mRNA的结合需要通过GTP激活进行额外的不可逆化学修饰(因此在动力学校正中称为“动力学”)。也就是说,当tRNA被激活时,它有第二次机会从mRNA上脱落。mRNA的解离速率并不取决tRNA,因此,活化配合物的解离速率也近似为rC和rI。活化速率ka是由GTP水解决定的,所以它不取决于底物是C还是i。在稳态条件下,流入将等于活化复合物的流出,所以

现在,结合公式4.43和4.40给出了这种激活的解离机制的选择性为

这表明了不可逆的GTP水解步骤如何放大结合第一步的选择性。这一过程被称为动力学校正,它提高了在生长中的肽链中添加氨基酸的正确率。
====================================================
-总结
蛋白质有许多不同的生物作用,包括催化、光的转导、ATP能量转化为力和运动、门控、调节、信号转导、氧的运输、DNA的螺旋和解决、DNA转录成RNA、其他蛋白质的合成等等。许多这些作用都是基本原理的简单结果,即当效应配体与蛋白质结合时,它会触发构象变化,调节蛋白质中不同的结合事件,这通常远离效应结合位点。而许多这些行为都需要耦合结合。结合性质和动力学通常可以用使用结合多项式的简单建模方式来描述如Hill、MWC和KNF模型。
第四章完
-------------------------------------------
欢迎点赞收藏转发!
下次见!