引言
大脑发育的关键分子机制在啮齿动物中已有所了解,但在灵长类动物中仍然不清楚,这限制了研究者对高级认知能力起源和功能障碍的理解。此外,在包括人类在内的灵长类动物中,关于轴突投射路径上的丘脑区域和皮层区域多样化的早期分子事件的信息匮乏,尤其是在神经干细胞生成神经元和胶质细胞的过程中。2023年10月12日Science在线发表了题为Molecular programs of regional specification and neural stem cell fate progression in macaque telencephalon的文章。
研究者利用单细胞转录组学研究方法,解剖了恒河猴产前大脑的多个区域,并对超过761,000个细胞进行了单细胞RNA测序(从神经发生前期到中期胶质发生时期)。该研究阐明了在恒河猴大脑皮层发育过程中神经干细胞(NSCs)身份进展和神经元、胶质特性的区域化分子机制,突出显示了早期事件作为与脑疾病相关的潜在风险因素。这一资源将进一步探索灵长类动物和人类大脑的发育、进化和疾病等方面。
结果
对于产前猕猴大脑颞叶细胞的时空转录特征表征
研究者对12个产前恒河猴颞叶中的多个潜在区域进行了单细胞RNA测序(scRNA-seq),共收集了82个样本,从神经发生前的胚胎第37天(E37)开始,直到神经胶质发生中期,即E110 (图1A)。在E37-E78期间,作者确定了节段发育核(GE)、前部(A)/额叶(FR)、背外侧(DL)/运动-体感(MS)、后部(P, 颞-枕)/枕叶(OC)以及潜在的颞叶壁。在E93和E110时,作者进一步划分了GE、潜在的前额叶(PFC)、主要运动区(M1C)、顶叶(Par)、主要视觉区(V1C)、岛叶(Ins)和颞叶(Temp)皮层壁等更精细的区域。经过严格的质量控制,作者得到了761,529个高质量的细胞。
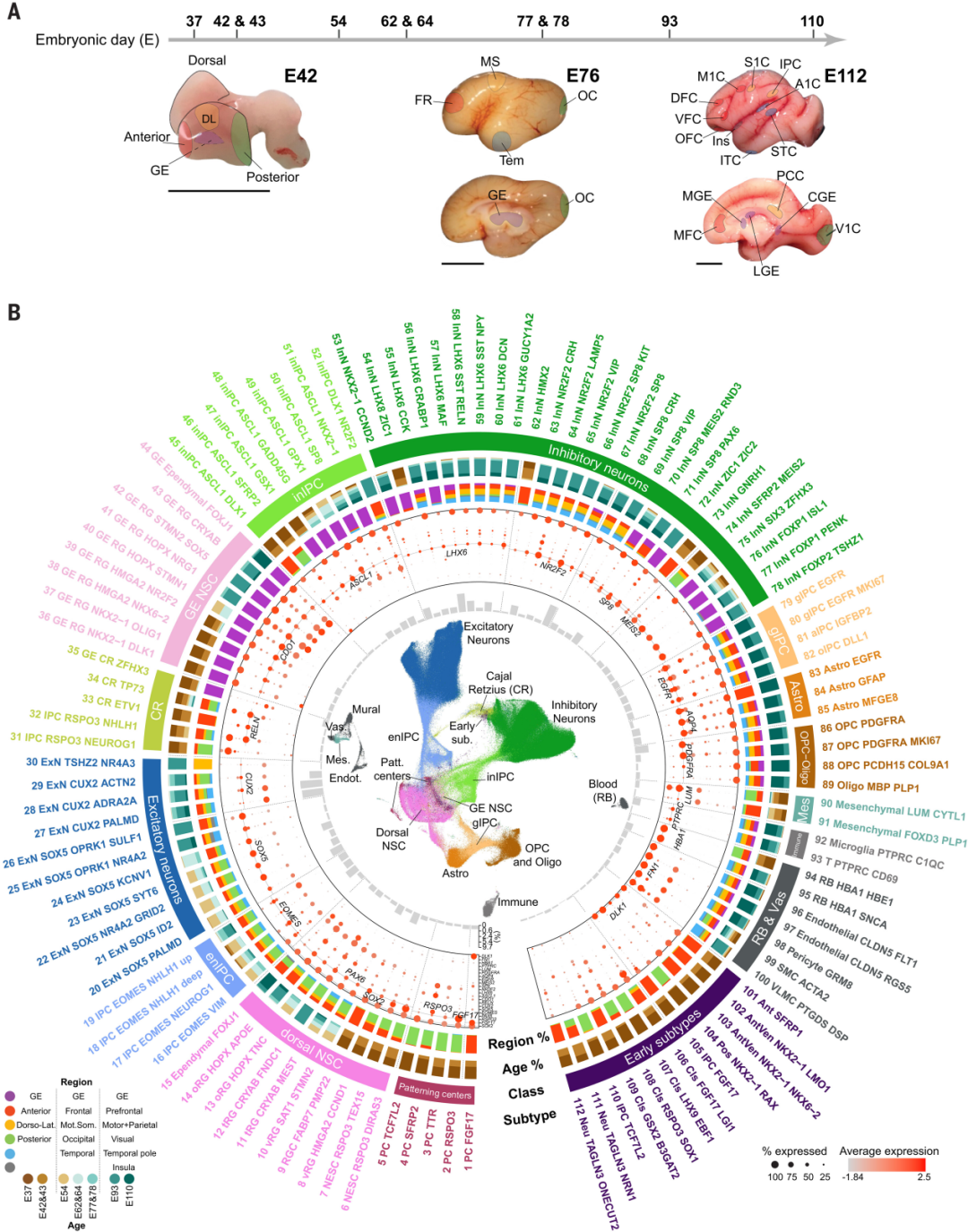
无监督聚类和mark gene分析确定了112个转录组学定义的细胞亚型,包括潜在颞叶定向中心的祖细胞;经典和腹侧神经干细胞分别经过兴奋性和抑制性神经发生谱系;胶质谱系;以及非神经细胞,这些细胞与样本的时空信息相对应,通过细胞上的统一流形逼近与投影(UMAP)布局呈现 (图1B)。
这个描绘了发展中猕猴颞叶单细胞时空转录动态的资源可在http://resources.sestanlab.org/devmacaquebrain上获得。
猕猴颞叶定向中心的转录组学特征
研究者确定了早期领域特异性的SOX2+/NES+神经外胚层祖细胞,代表着潜在的颞叶定向中心(PCs,也称为组织器)(图2, A到C)。在前部区域检测到的三个共聚类亚型表达FGF8/17/18、SP8、FOXG1、NKX2-1或SHH,代表着潜在的前神经脊带/前部定向中心 (RPC) (PC FGF17) 和前腹侧祖细胞 (AV) (AV NKX2-1/NKX6-2和AV NKX2-1/LMO1)。在GE中检测到的两个NKX2-1+亚型与AV祖细胞共聚类,可能代表着腹侧大脑组织器 。这些AV亚型中检测到了ZIC基因(ZIC1/3/4),与该区域中表达的AV定向基因形成了一个腹-背和前-后的梯度。在中后区域表达LMX1A、WNT (RSPO3、WNT8B)和BMP (BAMBI)信号分子的两个亚型可能代表背-尾状组织器(hem, PC RSPO3)和hem/脉络丛上皮(CPe, PC TTR)的两种状态,该区域的特征进一步通过ARX、FGFR3和LHX9的表达进行了描述 (图2, A到C)。其他后部亚型包括一个潜在的内丘脑界限带 (ZLI) (PC TCF7L2) 和一个抗hem (PC SFRP2)。这些细胞是暂时的,在E43后几乎无法检测到 (图2A),通过与小鼠和猕猴数据集的转录组比较进一步验证了它们的身份。
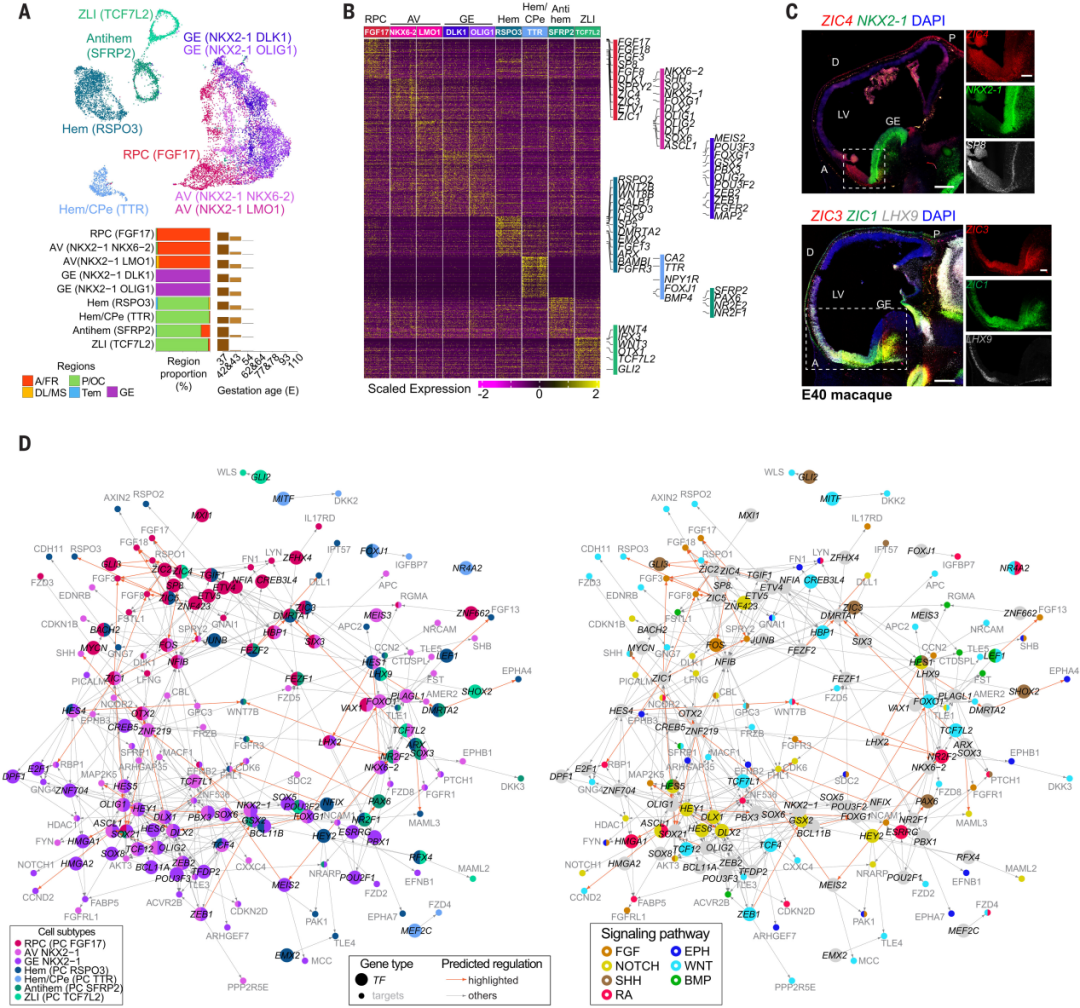
组织器分泌形态发生因子,从而在大脑皮层中引导基因表达梯度。通过整合基因motif富集和基因共表达信息,预测了连接转录因子(TFs)与其假定的靶基因的调控网络,包括在不同区域招募的信号分子成分(图2D)。RPC亚型可能通过上游的ZIC基因调控参与转录因子靶基因的调控,其中包括FGF(成纤维细胞生长因子)信号通路相关基因,如FGF3/8/17/18和SPRY2;而AV和GE祖细胞表现出重叠和亚型特异性的调控,包括将NKX2-1与SHH通过ZNF219连接起来。同样,后部组织器亚型显示出特定区域的调控,包括NFIX上游的Notch和FGF信号通路基因,以及区域共享的调控,如ARX和LHX9。同一信号通路的不同元素显示出不同的空间活化模式(图2D)。例如,BMP信号通路成员RGMA和FSTL1被招募到前-腹侧,而LEF1位于中部。这些数据共同证明了转录因子与信号分子的组合作用,协同调控组织器中心的模式形成作用。
最后,RNA速度推断了AV和皮层 HEM 祖细胞的潜在神经元谱系,分别生成 LHX8+ 和 ONECUT1+/ONECUT2+ 或 TP73+ 的 Cajal-Retzius 神经元。综上所述,这些数据突显了灵长类动物大脑端脑组织器活动的分子事件。
大脑端脑组织器与神经干细胞之间的信号交互
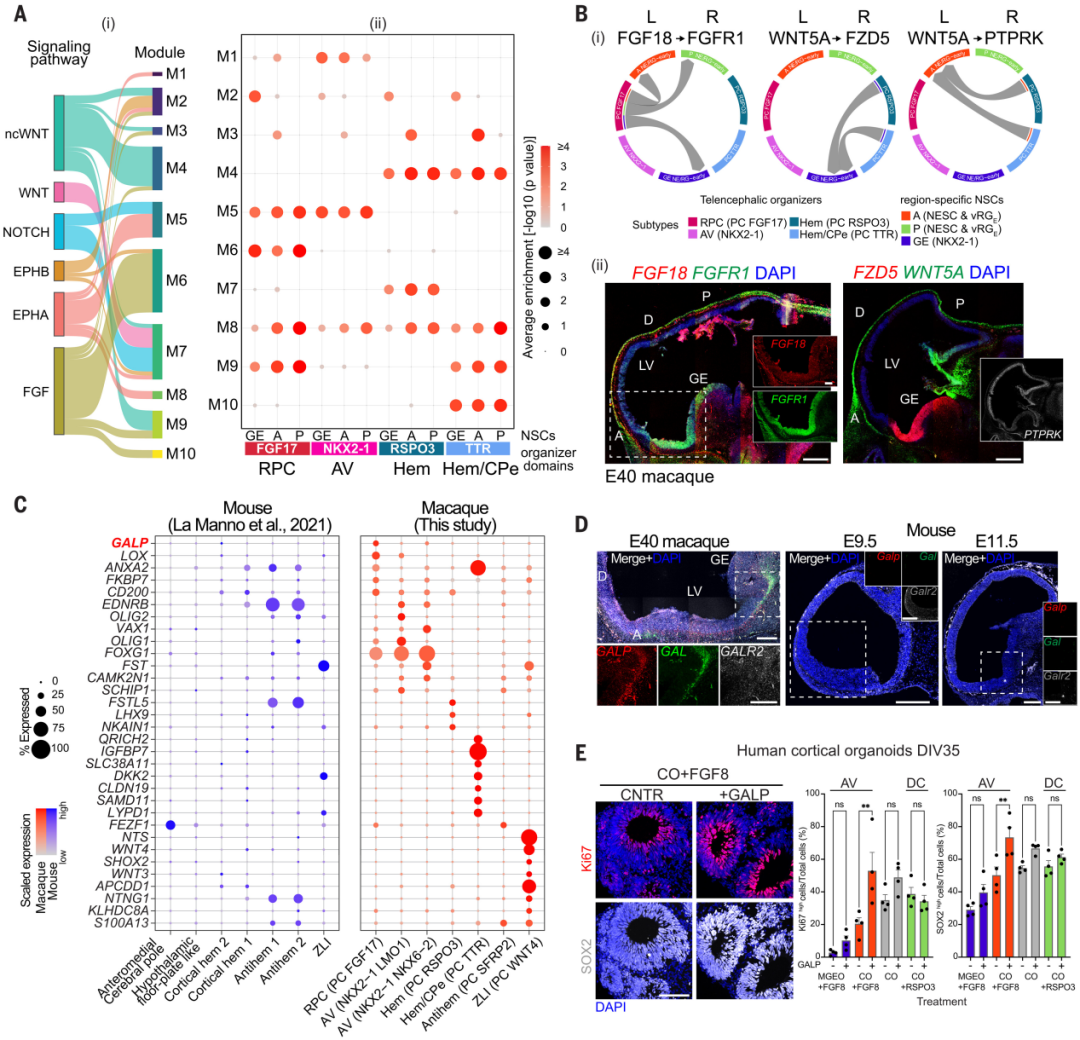
为了确定端脑放射状胶质细胞的区域特性是如何受到组织器的指导的,作者利用已注释的信号通路相关配体-受体(L-R)对,推测组织器亚型[RPC(FGF17+)、AV(NKX2-1+)、hem(RSPO3+)和hem/CPe(TTR+)]表达配体的细胞与特定区域的神经干细胞表达受体之间的细胞相互作用,并基于它们的相互作用模式(图3A、B),将它们聚类成模块(M1至M10)(图3A、B)。例如,M6主要由FGF信号通路的 L-R 对组成,其中包括FGF18-FGFR1和FGF18-FGFR3对,预测RPC细胞表达FGF18配体向表达FGFR1受体的前、后和腹侧神经干细胞发送信号,或选择性地向表达FGFR3的后部和腹侧神经干细胞发送信号。其他模块包括WNT(WNT5A-Frizzled受体FZD5,M2;WNT5A-WNT信号调节因子PTPRK,M3)和BMP信号相关的L-R对,预测了皮层HEM和区域特定神经干细胞之间的相互作用。RNAscope支持了预测的相互作用,描绘出了预期领域内的空间表达模式(图3B)。
这些数据强烈暗示大脑组织器有选择地向合适的神经干细胞发出信号。此外,结果显示猕猴中形态发生因子和配对受体的区域表达,支持了一个整合组织器和区域特定神经干细胞交互作用的信号编码假设。
潜在的灵长类动物偏向增殖信号
转录组比较表明,猕猴与小鼠大脑发育组织具有相似之处,也存在显著差异(图3C)。富集于猕猴发育组织的基因包括神经肽胆碱酯酶样肽(GALP),已知在啮齿动物成年时涉及下丘脑功能。作者发现,GALP在猕猴RPC祖细胞及其潜在后代谱系中表达。此外,它的表达在E43之后下降,表明其在早期阶段具有短暂的功能(图3C)。RNAscope分析验证了GALP及其与其相关的神经肽胆碱酯酶(GAL)在E40猴子端脑袋前区域的表达,而它们在小鼠E9.5和E11.5时期未被检测到(图3D)。GALP/GAL受体2(GALR2)在猴子中也很明显,但在小鼠端脑中较弱。
生成人类皮质(hCO)和中部GE(hMGEO)有机体,并在定型阶段调节RSPO3和FGF8信号以向背侧或前腹侧身份定向(29)。标准标记确认了它们的区域偏向;另外,研究者观察到hMGEO中GAL、GALP、ZIC4和SP8的表达比hCO更高,提示这些是前腹侧神经细胞的内在特征(图S7,A和B)。此外,将恒河猴皮层有机体(rmCO)暴露于GALP、GAL或两种配体中的任何一种,会增加NSC增殖并损害神经元分化(图S7,C和D)。
最后,外源性的GALP增加了具有前馈身份的hCO(hCO+FGF8)中的Ki67+和SOX2+细胞数量,而在前腹侧的hMGEO(hMGEO+FGF8)或背下的hCO(hCO+RSPO3)中没有增加(Fig.3E和图S7E)。此外,在E11.5小鼠胎儿的脑室内注射GALP配体,随后进行5-乙炔基2'-脱氧尿嘧啶(EdU)掺入,在E12.5相对于磷酸缓冲盐(PBS)注射对照组,在头骨中部背侧端脑中发现了更高比例的EdU+、Sox2+和Ki67+细胞,但在尾端和腹侧区域没有发现(图S8,A至C)。这些数据表明,GALP优先诱导具有前馈身份的大脑RG细胞增殖。
跨皮层区域NSC进程的转录差异
根据marker gene的表达,将皮层NSC区分为多个亚型,显示出不同的区域比例,并且它们的出现与发育年龄相关:两个神经上皮干细胞(NESC)亚型,两个早期(vRGE)和一个晚期(vRGL)脑室RG细胞亚型,两个截短RG(tRG)细胞亚型,一种室管膜亚型和两个外部RG(oRG)细胞亚型(图4A和B)。伪时间分析进一步定义了脑室NSC的进程,直到室管膜细胞,区分了oRG细胞谱系。与人类发育期脑单细胞RNA测序数据集的转录组比较验证了本研究中的注释,并确定了该项目数据集中整个半球发展的早期NSC状态。在分析的所有四个主要区域中,大部分亚型都存在;然而,在前方区域中选择性富集了一种早期RG细胞亚型(vRGE PMP22+),高表达CYP26A1和ZIC1/3/4(图4B)。
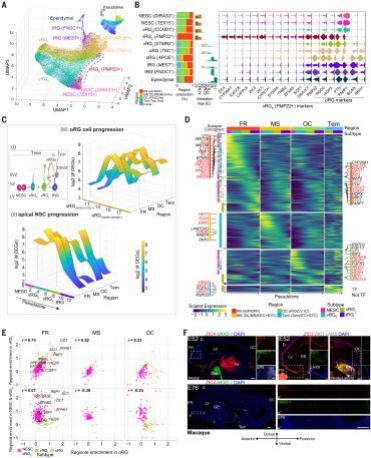
在区域共享的亚型中,例如在NESCs向vRG细胞转化期间,可检测到表达变化的染色质重塑因子(HMGA2和JARID2);在从tRG向室管膜细胞转化过程中发现了与纤毛相关的基因(FOXJ1);在oRG细胞发育过程中,细胞相互作用(neurexins)和血管生成(VEGFA)基因也有表达变化。因此,该分析确定了支配关键皮层干细胞状态转换的基因级联反应。
对于脑室区(VZ,也称脑室或顶部祖细胞)和亚脑室区(SVZ,也称基底祖细胞)的NSCs的不同表达基因(DEGs)进行的分析显示,在早期NSCs(NESCs和vRGE)和晚期oRG细胞中,区域多样性加强。在脑室NSC进展中,区域富集的基因表达级联反应在前/额叶细胞的早期阶段比较显著,包括转录因子(TFs)如ZNF219,ZIC1/2/3/4/5和SOX21,WNT成员(WNT7B,WNT8B),以及视黄酸(RA)信号分子(CYP26A1和RBP1)。在这些NSCs中,一些oRG标记基因(HOPX,PTN,FABP7和PMP22)得到富集,代表了早期前部种群的独特特征;但是在更成熟的状态中,它们的表达变得区域可比较。早期后部/枕叶区和颞区都显示出NR2F1/2、FGFR3、WNT(RSPO3)和Notch(HES5)信号成员的更高表达。
作者还发现,130个早期前/额叶和后部/枕叶NSCs的共享基因以及16种其他基因,包括神经肽PENK,其表达从早期前/额叶向晚期枕叶NSCs转移(fig. S10,D-G)。
在oRG细胞谱系中,跨区域的DEGs包括前部的RBP1;后部的MEF2C和NPY神经肽;以及颞部的BDNF受体NTRK2。因此,在顶部和基底NSC进展过程中,定义了时间调节的区域特异性基因表达模式。
最后,vRGL而不是早期NSCs(NESC和vRGE)与oRG细胞呈正相关,前部更加显著,只有少量的基因(例如RBP1、ZIC1和DCT)在所有前部NSC亚型中都有稳定的表达。这表明,顶部NSCs的区域特异性分子程序在早期阶段与oRG细胞不同,但稍后会传递给它们。然而,所有前部顶部NSCs和oRG细胞可能共享分子机制,包括RA信号响应。
猴脑组织的RNAscope验证了几个转录因子在E40、E52和E76的表达模式,例如ZIC4、SP8、NKX2-1、LHX9、FEZF1和NR2F1,它们在各自的区域随时间降低,而其他转录因子,例如ZIC1/3、MEIS2和PBX1,其表达增加,从前后向前轴扩散(图2C和4F)。因此,早期生成的梯度被发现是短暂的,在发育过程中改变其空间表达。总之,在灵长类脑的区域中,存在一系列顺序调节的基因,从NSCs开始沿着定义的区域特异性状态转换进行。
跨潜在皮层区域和区域的兴奋性神经元转录组多样性的分化
经无监督聚类和标记物 profiling 鉴定了兴奋性神经元系列中的不同亚型,从 EOMES+ 中间前体细胞(IPC)到深层(DL,SOX5+)和上层(UL,CUX2+)兴奋性神经元(图5A)。与成年猕猴 PFC 数据集整合,进一步描绘了这些轨迹,预测了胎儿神经元的成熟身份(图5B和图S11、A、C和D)。DL 神经元出现在E37-43,并在E54-64达到高峰,迅速分化为下板层 (L6B,from NR4A2+/GRID2+)、皮质丘脑通路 (L6CT,from SYT6+)和脑内皮层 (L6IT-1和L6IT-2,from OPRK1+) 亚型。UL 神经元系出现于 E77-78 并在 E93 得到增强,但其成熟进入成年细胞类型的分化并不明显,这表明需要额外的时间进行成熟。神经发生动态也因皮层区域而异。例如,枕叶UL神经元的出现比其他区域晚,但成熟速度更快。这些分析定义了支配灰质的细胞动态,其中不同的新皮质区域被基于其对应层的亚型所定义。
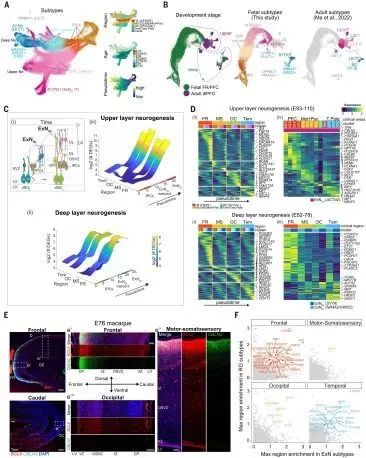
在兴奋性神经元假时序和分层聚类中,差异表达和AUC (曲线下面积) 分数分析突显了在晚期分化阶段区域和区域分化的增加,识别出更多的区域特异基因在额叶皮层 (图5C和D)。额叶 DL 神经元富集的基因包括原癌细胞粘附蛋白 (PCDH10/17),而 UL 神经元的区域差异则由 PFC 中的 RA 信号成员 (CYP26A1、CBLN2 和 MEIS2) 和 M1C 中的 BCL6 定义,如先前报道的 (10, 31) (图5D)。然而,在 IPCs 中也检测到非常显著的区域差异,这可能代表种子神经元分化的早期细胞自主事件。此外,虽然 DL 神经元亚型之间的区域特异性签名很大程度上重叠,但它们很少在 DL 和 UL 神经元之间共享,这表明在内外皮层层中建立区域身份的不同分子程序。RNAscope 在猕猴大脑上的应用进一步证实了在区域间存在差异表达模式,描绘了 BCL6 和 CBLN2 的前后梯度 (图5E)。最后,与年龄匹配的人类原代兴奋性神经元的转录组比较显示出人类偏向的特征,包括 RBP1,在背侧前额皮层中表达更明显。这些分析勾勒出了控制新皮质兴奋性神经元区域多样性的发育动态,突出了它们在成熟过程中身份的细化。
接下来,研究者探究了RG 细胞的原型图是否与兴奋性神经元的区域特异性相关。通过交叉分析早期 RG 细胞和神经元的区域特异基因,发现这些基因在整个谱系发展过程中都有表达。这些包括 HOPX、CYP26A1 和 RBP1 在前额、NR2F1 和 RSPO3 在枕叶以及 NR2F2 和 CYP26B1 在颞区,可能定义了神经细胞在发育过程中的区域身份 (图5F)。此外,单个神经元的 RNA-seq 分析,跨越猴子区域性皮质 NSCs 在体外分化 ,鉴定了许多表达在神经元中的区域特异基因,这些基因在 vivo 中的兴奋性神经元中也存在,包括 RA 信号成分 (RBP1、CYP26A1、BRINP1 和 CBLN2) 和突触相关基因 (LFRN2 和 CAMKV) 。由于这些体外神经元缺乏其他脑区的连接输入,这些确定的区域特异基因可能反映了神经元区域分化的内在事件,并且在前额皮层中更为突出。总之,这些数据揭示了早期和晚期细胞自主机制对新皮质区域间神经元的多样性的潜在贡献。
相比之下,抑制性神经元作为分布在各个区域和年龄的亚型,例如 MGE (LHX6+) 和 CGE (NR2F2+ 和/或 SP8+) 派生的亚型,显示出有限的皮层区域差异。在 LHX6+ 抑制性神经元中表达的特定区域基因与 NR2F2+/SP8+ 亚型中表达的基因完全相似,反之亦然,表明抑制性神经元之间地区身份的转录重叠,并暗示后期信号可能最终促进它们在皮层中的进一步多样化。
神经发育过程中的空间转录组差异
接下来作者关注神经胶质形成的轨迹。通过无监督聚类、细胞轨迹重建推断和拟时序分析,鉴别出了晚期胶质母细胞亚型,包括vRGL、tRG和oRG细胞,它们分化为兴奋性神经元或EGFR高表达的胶质中间前体细胞(gIPCs),后者分化为星形胶质细胞或少突胶质细胞(图6A、B)。与多个胎儿和成年人类、猕猴和小鼠数据集的比较分析确认了该注释结果,并区分出了假定层间(GFAP+)和原浆质(MFGE8+和EGFR+)亚型的星形胶质细胞,表明星形胶质的成人身份在灵长类动物的中期胚胎阶段出现。

为了定义RG细胞从产生神经元潜能向产生胶质潜能转变的转录组程序,作者确定了每个谱系分支上表达的排名靠前的基因(图6C)。已知的星形胶质命运调控因子如染色质重塑因子HMGN3和HMGB2以及转录因子PAX6、HES1和SOX2/3/6由晚期RG细胞表达;OLIG1/2和ASCL1由gIPCs表达;而少突胶质细胞则表达SOX10和NKX2-2,而星形胶质细胞表达了许多RG基因。这些数据揭示了在灵长类脑皮质发生过程中从神经发生到胶质发生的时间上有序的基因组合。
差异表达基因分析表明,胶质细胞的差异性小于兴奋性神经元。然而,星形胶质细胞在E110时显示出更多的差异表达基因,包括转录因子,相比之下gIPCs和少突胶质细胞的差异表达基因较少,并表达区域特异性基因,例如前额叶皮质中的SLC35F1和颞叶皮质中的STAT3,表明它们具有不同的空间分子特征(图6D、E)。这些结果表明,星形胶质细胞在后期分化过程中的转录组变异在不同脑区更为显著。
神经元和非神经元细胞之间的左右配对分析预测到最多的潜在相互作用,其中最明显的区域差异是在内皮细胞和星形胶质IPC或星形胶质细胞之间(图6F)。内皮细胞在不同脑区的转录变异较小,这表明星形胶质细胞可能对其信号有不同的响应。研究者还确定了多种信号通路的左右配对,在时间和空间上都显示出变异。Notch和Midkine (MDK)信号相关的左右配对在内皮细胞中显示出相似的配体(JAG1/2、DLL4和MDK),而在星形胶质细胞中找到了受体(NOTCH1/2/3和SDC2以及ITGB1)的变异(图6G)。这些数据表明,星形胶质细胞具有区域特异性的反应内皮细胞信号的能力,这可能是其在不同脑区的转录组变异中的因素。
早期端脑发育中与疾病风险基因的时空表达
皮质发育的异常与神经精神类疾病有关,然而,关于这些风险相关基因在早期端脑神经干细胞中的功能知之甚少。研究者整理了与多种神经精神类疾病、神经退行性疾病以及脑癌相关的基因列表。表达富集分析显示多种神经精神类疾病的风险基因在兴奋性和抑制性神经元中显著表达,而胶质母细胞与胶质前体细胞中高表达脑胶质瘤相关基因也得到了证实(18, 37-40)(图7A)。