什麼是「垃圾DNA」? | GeneOnline News
垃圾dna与转座子
基因组重复序列分类 转座子 - 组学大讲堂问答社区 (omicsclass.com)
(122条消息) 说说基因组的垃圾DNA-Transposable elements_msw521sg的博客-CSDN博客
Cell:转座子“跳跃”过程调控机制 - 生物研究专区 - 生物谷 (bioon.com)
分类
转座元件,是一种跳跃基因,是指一段DNA序列由基因组的一个位置跳跃到另一个位置,由Barbara McClintock教授在大约五十年前发现。转座子主要包含两种类型:
一类转座子: retrotransposons,通过RNA的媒介作用,通过RNA的反转录获得DNA,从而转移到其他基因组位置。
二类转座子:DNA transposons
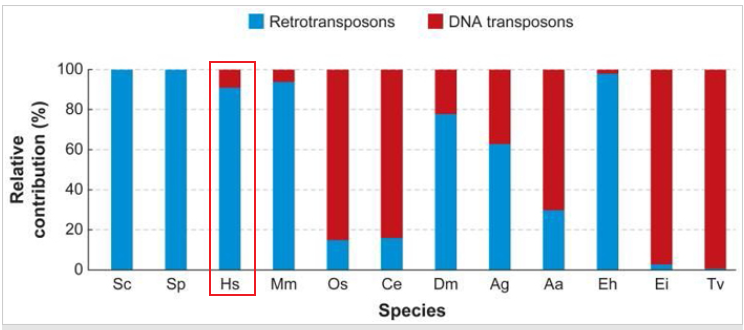
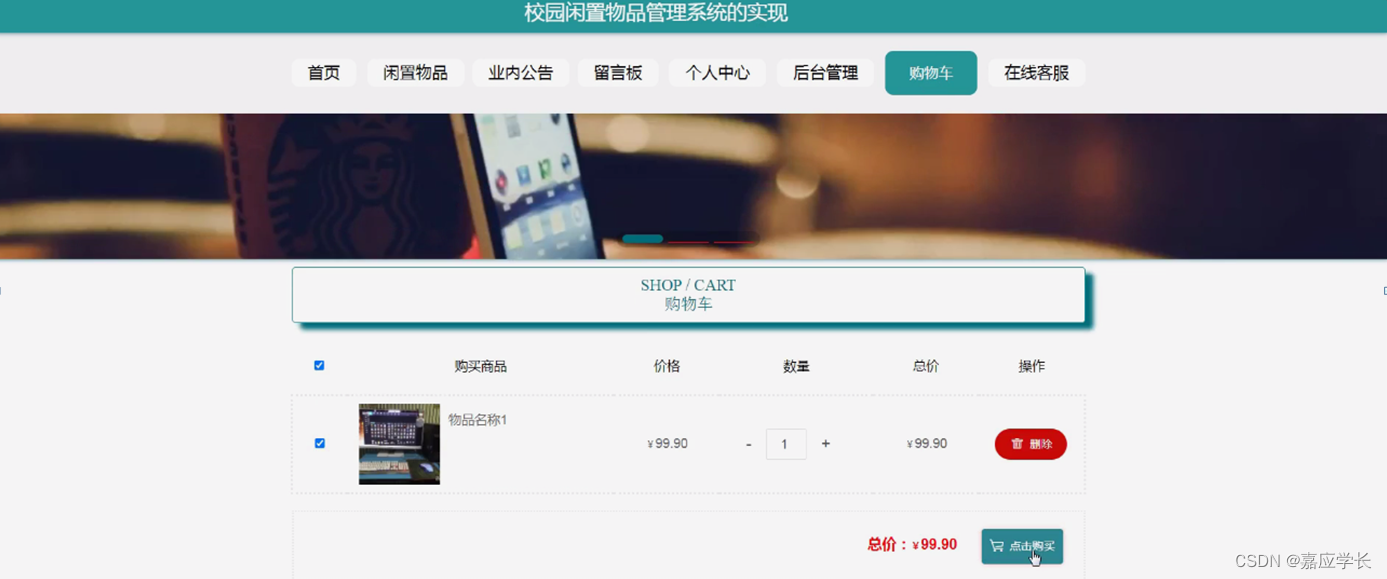
转座子的分布.png
如图所示,人类中90%都是一类转座子,主要一类逆转录酶来讲RNA转换为DNA,然后插入到目的基因组的位置。目前有主要有两种类型的一类转座子:1.LTR retrotransposons,long terminal repeats,双末端都是长的重复序列;2.non-LTR TEs,双末端缺乏重复序列。LINE1(L1)和 Alu基因都是 non-LTR;L1元件通常是6 kilobases的长度;相比较而言,Alu元件平均只有几百核苷酸,因此他们是短穿插转座元件(short interspersed transposable element:SINE),Alu特别多产,起源于灵长类,并在较短的时间内扩展到人类每个细胞约一百万个拷贝;L1在人类中也非常普遍,虽然拷贝数量不像Alu那么多,但是由于其长度较长因此在人类基因组中占大约15%-17% (Kazazian, H. H., & Moran, J. V. The impact of L1 retrotransposons on the human genome. Nature Genetics 19, 19–24 (1998) ; Slotkin, R. K., & Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nature Reviews Genetics 8, 272–285 (2007))。这些non-LTR TEs是唯一活跃的转座子类别。 LTR retrotransposons 和DNA转座子只是古老的基因组遗物,不能进行基因组跳跃。
结构
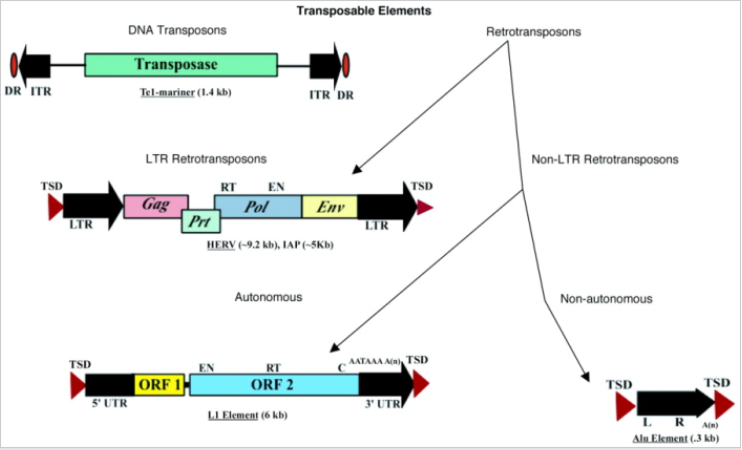
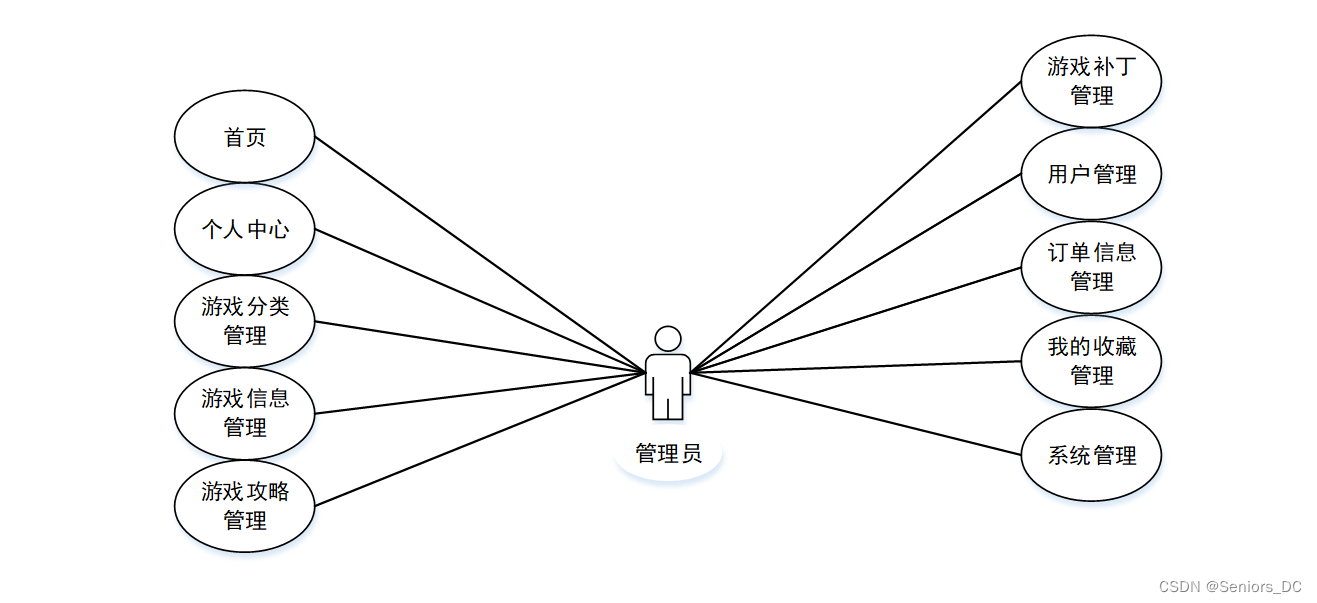
image.png
逆转座子 Retrotransposons 根据是否具有编码逆转座所需的ORF分为自主和非自主类。普通的自主类Retrotransposons 分为(i) LTRs or (ii) non-LTRs。图上所示的LTR retrotransposons的例子是人类内源性逆转录病毒(HERV);L1是 non-LTRs的例子,包含一个5' UTR有一个内部启动子,两个开放阅读框,一个3' UTR,和一个polyA信号加上polyA尾。L1的两侧通常是7-20bp的靶向位点重复(TSDs)。Alu元件是 nonautonomous retrotransposon的一个例子,包含两个相似的单体,左和右,结尾一个polyA尾。图中括号内给出了元件全长的大小。
图中的一些缩写:
- inverted terminal repeats (ITRs) 反向末端重复
- open reading frame (ORF)开放阅读框
- direct repeats(DRs)直接重复
- human endogenous retroviruses (HERV) 人类内源性逆转录病毒
- reverse transcriptase (RT) and endonuclease (EN) domains逆转录酶(RT)和核酸内切酶(EN)域
- 5'-untranslated region (5' UTR)
- target site duplications (TSDs) 靶向位点重复
- poly(A) tail (An) polyA尾
图片来自:American Association for the Advancement of Science Kazasian, H. H. Mobile elements: drivers of genome evolution. Science 303, 1626–1632 (2004).
自主和非自主的Transposons
1类和2类TE可以是自治的也可以是非自治的。 自治TE可以自行移动,而非自治元素需要存在其他TE才能移动。这是因为非自治元件缺少转座所需的转座酶或逆转录酶基因,因此它们必须从另一个元件“借来”这些蛋白质才能移动。例如,Ac元件是自主的,因为它们可以自行移动,而Ds元件是非自主的,因为它们需要Ac的存在才能进行转置。
基因除了跳跃还做了什么
人类基因组中大约一半由TE组成,其中很大一部分是L1和Alu逆转座子,这一事实提出了一个重要的问题:除了跳跃外,所有这些跳跃基因还做什么? 转座子的大部分功能取决于它的着陆位置。 如将L1插入VIII因子基因引起血友病时发现,落在基因内部可导致突变(Kazazian, H. H., et al. Haemophilia A resulting from de novo insertion of L1 sequences represents a novel mechanism for mutation in man. Nature 332, 164–166 (1988) )。 同样,几年后,研究人员在同一个人的结肠癌细胞的APC基因中发现了L1,但在健康细胞的APC基因中未发现L1。 这证实了L1在哺乳动物的体细胞中转座,并且该元素可能在疾病发展中起因果作用(Miki, Y., et al. Disruption of the APC gene by a retrotransposal insertion of L1 sequence in colon cancer. Cancer Research 52, 643–645 (1992))。
转座子的沉默
与L1相反,大多数TE似乎是沉默的-换句话说,这些元件不会产生表型效应,也不会在基因组中主动移动。至少这是普遍的科学共识。 一些沉默的TE是失活的,因为它们的突变会影响其从一个染色体位置移动到另一个位置的能力。 其他分子则完好无损且能够移动,但通过表观遗传防御机制(例如DNA甲基化,染色质重塑和miRNA)保持失活。 例如,在染色质重塑中,对染色质蛋白的化学修饰导致染色质在基因组的某些区域变得狭窄,以致于这些区域中的基因和TE被沉默,因为转录酶根本无法访问它们。因为转座子的运动可能具有破坏性,所以人类基因组中的大多数转座子序列都是沉默的也就不足为奇了,尽管TE盛行,但该基因组仍保持相对稳定。实际上,研究人员认为,在由L1相关序列编码的人类基因组的17%中,仅剩余约100个活性L1元素。此外,研究表明,即使是少数剩余的活性转座子,也可以通过表观遗传沉默以外的多种方式来抑制跳跃。
转座子不并总是具有破坏作用
并非所有的转座子跳跃都会导致有害作用。实际上,转座子可通过促进基因组序列的易位,外显子的改组和双链断裂的修复来驱动基因组的进化。插入和转座也可以改变基因调控区和表型。转座子增加遗传多样性的能力,以及基因组抑制大多数TE活性的能力,导致平衡,使转座因子成为所有携带这些序列的生物中进化和基因调控的重要组成部分。