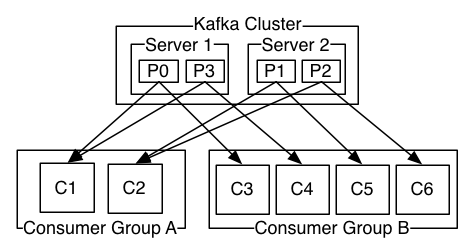
作者:非妃是公主
专栏:《智能优化算法》
博客地址:https://blog.csdn.net/myf_666
个性签:顺境不惰,逆境不馁,以心制境,万事可成。——曾国藩
文章目录
- 专栏推荐
- 序
- 一、生物进化
- 二、遗传算法原理
- 三、遗传算法的优点
- 四、算法具体流程
- 五、仿真实例:求解函数最大值
- 1. 题目
- 2. 分析
- 3. matlab求解
- 4. 求解结果及分析
- 六、遗传算法的一些改进方向
- the end……

专栏推荐
| 专栏名称 | 专栏地址 |
|---|---|
| 软件工程 | 专栏——软件工程 |
| 计算机图形学 | 专栏——计算机图形学 |
| 操作系统 | 专栏——操作系统 |
| 软件测试 | 专栏——软件测试 |
| 机器学习 | 专栏——机器学习 |
| 数据库 | 专栏——数据库 |
| 算法 | 专栏——算法 |
序
遗传算法也是一种效果很棒的智能优化算法,它主要借鉴了自然界中生物进化的思想,采用优胜劣汰的策略,利用选择、交叉、变异三个过程,对解进行不断的迭代,最终趋向于最优解。
一、生物进化
谈到遗传算法,不得不谈生物进化,因为遗传算法就是模仿生物繁殖后代的思想演变而来的,可以说是一种仿生学的算法。
自然选择学说认为,适者生存,生物想要存活下去,就必须进行生存斗争。生存斗争包括种内、种间以及生物和环境之间的斗争三个方面。
在斗争过程中,具有有利变异的个体容易存活下来,并且有更多的机会将有利变异传递给后代;具有不利变异的个体就容易被淘汰,产生后代的机会也就少得多。因此,凡是在生存斗争中获胜的个体都是对环境适应性比较强的个体。达尔文把这种在生存斗争中适者生存,不适者淘汰的过程叫做自然选择。1
二、遗传算法原理
而遗传算法就是借鉴了上述生物进化中的优化策略,对一个目标问题进行优化的。
具体地说,它将问题的解抽象成一个编码,许多个编码构成了一个种群,每个编码都对应得到目标问题的一个解,其中根据可行解的目标函数值的不同,定义这个个体的强弱。越强的个体越容易生存下来(选择),进行后代的繁衍(交叉、变异),越弱的个体就优先被淘汰。
| 生物进化中的术语 | 遗传算法中的术语 |
|---|---|
| 群体 | 可行解集 |
| 个体 | 可行解 |
| 染色体 | 可行解的编码 |
| 基因 | 可行解编码中的一位(1个bit) |
| 适应度 | 评价函数值(越高,代表解越好) |
| 选择 | 选择操作(比如根据概率进行轮盘赌选择可行解) |
| 交叉 | 交叉操作(编码的单点交叉、多点交叉) |
| 变异 | 变异操作(编码的按位取反) |
三、遗传算法的优点
- 以决策变量的编码作为运算对象。这种编码的方式对于计算机来求解十分方便,具有独特的优越性。
- 遗传算法以目标函数值作为搜索信息,避免了求导。在很多问题中,求导是无法做到的或者很困难做到的。
- 选择、交叉、变异操作包括了很多群体信息。这些信息避免搜索了许多的点,相当于具有一种隐含的并行性。
- 遗传算法基于概率。随着过程的进化,参数对搜索结果的影响很小很小。
- 具有自组织、自适应、自学习等特性。
四、算法具体流程
1. 整体流程
- 初始化;
- 个体评价;
- 选择运算;
- 交叉运算;
- 变异运算;
- 终止条件判断。
流程图如下:
2. 细节处理
Ⅰ. 群体规模 N P NP NP
群体规模影响遗传优化的最终结果以及遗传算法的执行效率。一般 NP取10~200。
- N P NP NP 太小时,遗传优化效果(相比于最优解准确度)一般不会太好。
- 较大的群体规模可以减小陷入局部最优解的概率,但意味着较大的计算复杂度。
Ⅱ. 交叉概率 P c P_c Pc
交叉概率 P c P_c Pc控制着交叉操作的频率,较大的频率可以曾倩遗传算法开辟新的搜索区域的能力。
- 如果太大,容易生成不好的后代,收敛性差;
- 如果太小,搜索能力差。
Ⅲ. 变异概率 P m P_m Pm
保持群体多样性。一般去0.001~0.1
- 太大,容易陷入随机搜索,导致重要基因丢失。
- 太小,没有效果。
3. 遗传运算终止代数 G G G
一般视具体问题而定,取值可在 100~1000 之间。
- 太大,浪费算力,种群衰退。
- 太小,优化过程没有收敛。
五、仿真实例:求解函数最大值
1. 题目
用标准遗传算法求函数 f ( x ) = x + 10 s i n ( 5 x + 7 c o s ( 4 x ) f( x ) =x+10sin(5x +7cos(4x) f(x)=x+10sin(5x+7cos(4x)的最大值,其中的取值范围为 [ 0 , 10 ] [0,10] [0,10]。
2. 分析
通过matlab作图可知,该函数存在多个局部极值,如下图所示:

- 初始化种群NP=50,染色体编码长度L=20,最大进化代数G=50,交叉概率为 P c = 0.8 P_c=0.8 Pc=0.8,变异概率为 P m = 0.1 P_m=0.1 Pm=0.1。
- 产生初始种群,将二进制编码转换成十进制,计算个体适应度值,并进行归一化;采用基于轮盘赌的选择操作、基于概率的交叉和变异操作产生新的种群,并把历代的最优个体保留在新种群中,进行下一步的遗传操作。
- 判断是否满足终止条件。满足,结束;不满足,继续迭代。
3. matlab求解
matlab求解代码如下,其中主要思路及较为复杂的点,已在注释中说明。
%%%%%%%%%%%%%%%%%%%%标准遗传算法求函数极值%%%%%%%%%%%%%%%%%%%
%%%%%%%%%%%%%%%%%%%%%%%%初始化参数%%%%%%%%%%%%%%%%%%%%%%%%%%
clear all; %清除所有变量
close all; %清图
clc; %清屏
NP=50; %种群数量
L=20; %二进制数串长度
Pc=0.8; %交叉率
Pm=0.1; %变异率
G=100; %最大遗传代数
Xs=10; %上限
Xx=0; %下限
f=randi([0,1],NP,L); %随机获得初始种群
finalMax=0;
finalXBest=0;
%%%%%%%%%%%%%%%%%%%%%%%%%遗传算法循环%%%%%%%%%%%%%%%%%%%%%%%%
for k=1:G % 跌代的总代数
%%%%%%%%%%%%将二进制解码为定义域范围内十进制%%%%%%%%%%%%%%
for i=1:NP % 遍历所有个体
U=f(i,:); % 将第 i 个个体读取出来
m=0; % 存储的二进制对应的十进制数字
for j=1:L % 按位把二进制转化为十进制
m=U(j)*2^(j-1)+m;
end
x(i)=Xx+m*(Xs-Xx)/(2^L-1); % 把十进制数字m转换到对应的定义域内
Fit(i)= func1(x(i)); % 计算适应度函数
end
maxFit=max(Fit); %最大值
minFit=min(Fit); %最小值
rr=find(Fit==maxFit); %找出最优个体的下标索引
fBest=f(rr(1,1),:); %当代最优个体的二进制编码
xBest=x(rr(1,1)); %当代最优个体对应的十进制值
if maxFit>finalMax
finalMax=maxFit;
finalXBest=xBest;
end
Fit=(Fit-minFit)/(maxFit-minFit); %归一化适应度值
%%%%%%%%%%%%%%%%%%基于轮盘赌的选择操作%%%%%%%%%%%%%%%%%%%
sum_Fit=sum(Fit); %对所有个体的适应度进行一个求和
fitvalue=Fit./sum_Fit; %让所有个体加起来适应度值为1,便于进行概率计算
fitvalue=cumsum(fitvalue); %consum是累加当前索引及以前的索引对应的值,比如[1 2 3],consume后返回[1 3 6]
ms=sort(rand(NP,1)); %生成一个NP * 1的向量,并从小到大排序,轮盘赌的关键操作。(均匀分布)
fiti=1;
newi=1;
while newi<=NP % 这个while循环是轮盘赌的精髓,如果适应度(存活概率)越大的个体,随机数越容易连续着比他小
if (ms(newi))<fitvalue(fiti) % 优胜个体,多复制几个
nf(newi,:)=f(fiti,:); % 复制
newi=newi+1; % 新个体计数+1
else % 劣汰个体,少复制或者干脆直接跳过去
fiti=fiti+1;
end
end
%%%%%%%%%%%%%%%%%%%%%%基于概率的交叉操作%%%%%%%%%%%%%%%%%%
for i=1:2:NP % 此处步长为2,主要是相邻两个个体进行交叉
p=rand; % 生成一个随机数
if p<Pc % 以Pc的概率进行交叉
q=randi([0,1],1,L);% 随机生成一个交叉序列,0位不变异,1位变异
for j=1:L
if q(j)==1 % 交叉的基因,不交叉的基因直接跳过
temp=nf(i+1,j);
nf(i+1,j)=nf(i,j);
nf(i,j)=temp;
end
end
end
end
%%%%%%%%%%%%%%%%%%%基于概率的变异操作%%%%%%%%%%%%%%%%%%%%%%%
i=1;
while i<=round(NP*Pm)
h=randi([1,NP],1,1); %随机选取一个需要变异的个体(染色体)
for j=1:round(L*Pm) %Pm是变异概率,L * Pm四舍五入得到变异的基因数量
g=randi([1,L],1,1); %随机需要变异的基因数
nf(h,g)=~nf(h,g);
end
i=i+1;
end
f=nf; %更新种群
%由于轮盘赌选择的时候,在前面的个体更容易被选中,
f(1,:)=fBest; %如果最优个体在后面,则不一定被选中,这步操作是要保留最优个体(二进制)在新种群中
trace(k)=maxFit; %历代最优适应度
end
xBest %最后一代最优个体
finalXBest %最优个体
figure
plot(trace)
xlabel('迭代次数')
ylabel('目标函数值')
title('适应度进化曲线')
其中目标函数(适应度函数)的定义如下:
%%%%%%%%%%%%%%%%%%%%%%%%%适应度函数%%%%%%%%%%%%%%%%%%%%%%%%%%%%
function result=func1(x)
fit= x+10*sin(5*x)+7*cos(4*x);
result=fit;
4. 求解结果及分析
可以看到,遗传算法的收敛速度较快,如下:

同时,遗传算法收敛性很棒的特点,经过多次测试,最后一代的最优个体就是所有代中的最优个体,即:在该问题中种群没有发生退化,导致目标函数出现震荡的现象。

六、遗传算法的一些改进方向
在现有文献中,对遗传算法的改进主要集中在以下方面:编码机制(如何编码(转化问题))、选择策略(怎么去选择?)、交叉算子(怎么交叉?)、变异算子(怎么变异)、特殊算子和参数设计。
每一个遗传算法的应用都要涉及到上述问题,上面的案例代码中包含了这些方面一种解决办法,当然,后续的改进还有许多。
此外,还存在着和差分进化算法、免疫算法、蚁群算法等结合起来的混合遗传算法,可以综合遗传算法和其它算法的优点,提高效率和求解质量。2
the end……
遗传算法到这里就要结束啦~~到此既是缘分,欢迎您的点赞、评论、收藏!关注我,不迷路,我们下期再见!!
😘😘😘 我是Cherries,一位计算机科班在校大学生,写博客用来记录自己平时的所思所想!
💞💞💞 内容繁杂,又才疏学浅,难免存在错误,欢迎各位大佬的批评指正!
👋👋👋 我们相互交流,共同进步!
注:本文由
非妃是公主发布于https://blog.csdn.net/myf_666,转载请务必标明原文链接:https://blog.csdn.net/myf_666/article/details/129166215
百度百科——自然选择 ↩︎
包子阳,智能优化算法及其matlab实例.电子工业出版社. ↩︎