导读
尤里卡效应(Eureka effect;灵光一现)是指突然解决问题的常见经验。在这里,本文以一种模式识别范式来研究这种效应,该范式要求在格式塔规则和先验知识的基础上分割复杂场景和识别物体。受试者必须在模糊的场景中检测物体,并通过手动反应来指示识别。采用高密度脑电图(EEG)记录评估神经动力学。结果显示,神经动力学在谱分布、相干性、锁相性和分形维数方面发生了显著变化。尤里卡效应与alpha和theta波段振荡的相干性增加有关,这些波段广泛分布在大脑皮层的覆盖区域,主要分布在右半球。这种相干性的增加与顶叶和中央区域的beta功率下降以及额叶和枕叶区域的alpha功率下降有关。此外,右侧半球的分形维数降低。本研究认为,尤里卡效应需要涉及工作记忆、创造性思维和注意控制的皮层区域的合作。
前言
神经系统需要能够区分与寻找计算问题的解决方案相关的活动模式和代表结果的活动模式。为了终止进一步的搜索,最终将结果转化为行动,为了允许突触可塑性来存储结果,为了允许启动新的搜索过程,以及允许结果进入意识,这种区分是必要的。人类将解决方案状态视为一种奖励,并且能够判断特定结果的可靠性。突然找到一个感性问题的解决方案,理解一个以前无法理解的概念或者解决一个难题,都是令人愉快的。这种从寻找解决方案到找到解决方案的突然转变被称为尤里卡效应。尤里卡经历的一些特征可能也是高等哺乳动物所共有的。
然而,人们对尤里卡效应的神经元基础知之甚少。其中一个原因是很难预测尤里卡效应在什么情况下以及何时发生。对尤里卡效应的研究存在一些方法论上的问题。首先,需要以高时间分辨率进行测量,因为尤里卡效应发生的时刻通常是不可预测的。其次,神经生理学研究需要足够数量的可比试次进行分析。
本研究采用黑白Mooney图像作为诱发尤里卡效应的范式。Mooney图像最有趣的特征是它们最初很难解释,但最终可以导致对物体的稳定感知。这种感知也可以通过线索来促进,线索通常由Mooney图像的原始图像(彩色或灰度图像)组成。Mooney图像作为测试材料有几个优点。首先,它们允许精确操作亮度、对比度、大小和像素等参数。其次,Mooney图像允许将自然物体与复杂背景相结合。这允许诱发强烈的尤里卡效应和寻找神经元关联。为了最大限度地出现尤里卡体验,本研究开发了一种生成和微调Mooney图像的方法,能够可靠且可重复地唤起尤里卡体验。
来自Mooney图像的模糊输入如何转化为明确的感知解决方案?建议使用存储在记忆中的先验经验来调节外围处理,以促进场景分割、感知分组和识别。贝叶斯感知假说认为,由自上而下的过程提供的内部信息与感官证据相匹配。因此,假设大脑执行概率推断,可以优化感官信息,以最大限度地减少内部和外部信息之间的不匹配。此功能可以解释为知觉闭合,它包括不完整感觉证据的完成以及将不同组成部分正确结合成一个连贯的知觉。有一种观点认为,这是通过将分布式神经元瞬时关联成一个连贯活跃的集合来实现的。
假设这些集合的特征是增强了神经元活动的一致性,那么尤里卡效应是否与神经元动力学的变化相关,如果是的话,那么尤里卡效应又是如何与神经元动力学的变化相关的。本研究假设解决方案状态可能与增强的神经元活动的一致性有关,因为它们可能是分布式计算结果成功整合的结果。为了验证这一假设,本研究采用Mooney刺激以诱发尤里卡效应,使用高密度EEG记录捕获神经活动,然后研究这些神经元反应在尤里卡效应期间是如何调节的。人脑是一个高度复杂和动态的系统,表现出广泛的状态转换。鉴于此,复杂性理论中使用的指标被认为是这些状态转换的宝贵指标,并可能为大脑功能的潜在机制提供重要的见解。近年来,非线性方法因其能够表征健康和病理性大脑活动的不同状态而引起了人们的极大兴趣。其中一个常用的指标是分形维数,它是衡量时间序列复杂程度的指标,现已被用于检测大脑在各种过程中的状态变化。例如,它已被用于区分不同的睡眠阶段,识别与癫痫发作相关的大脑活动变化,并作为各种疾病的潜在生物标志物,如自闭症、抑郁症和阿尔茨海默症。基于此,本研究不仅测量了振荡活动的功率和相干性,还测量了活动向量的分形维数,以进一步表征与尤里卡效应相关的状态转换。
材料和方法
图像的选择
用于构建刺激的原始图像来自Caltech-256数据集和Flickr。本研究的初始数据库中包含3万多张图像。然后,根据以下标准手动选择图像进行进一步测试。(i)熟悉度:本研究选择了众所周知的物体,如动物、植物、家具、工具、仪器、车辆等。(ii)自然复杂背景:图像取自自然场景,目标物体被嵌入在复杂背景中,这可能使识别变得困难。
处理图像的过程
首先将原始图像转换为256级灰度图像。由于灰度像素的分布范围大且非正态分布,并且亮度和对比度也不同,因此需要进行调整。通过直方图均衡化将亮度和对比度等同,得到均匀的对比度分布。
为了生成具有不同衰减程度的Mooney图像,将上述方法处理后的灰度图像进行频域高斯滤波。降低滤波器的截止频率会增加识别图像中物体的难度。滤波器的生成分为三个处理步骤:①通过傅里叶变换将原始图像f(x,y)的空间域变换到频域F(u,v);②用一组截止频率对F(u,v)进行低通滤波,得到一个频带通过表示G(u,v);③该表示G(u,v)通过傅里叶逆变换变换回空间域g(x,y)(即模糊图像)。为了获得具有不同衰减程度的图像,低通滤波器在10~80Hz(间隔为10Hz)的截止频率范围内变化。然后以图像的中值灰度作为黑白像素的分割线,得到两色调Mooney图像。随后,测试了不同截止频率下的Mooney图像的可识别性。本研究选择20Hz的截止频率来诱发尤里卡效应,因为它确保了从搜索到解决方案的最可靠转换。
参与者
在确定截止频率的预实验中,共纳入10名受试者,这些受试者均未参加主实验。另有29名健康受试者(年龄为25.2±4.0岁,16名男性,13名女性)参加了主实验,包括行为评估和EEG记录。这些受试者均未参加预实验。有4名受试者的数据因正确反应试次不足而被排除,3名受试者的脑电数据由于伪迹排除导致有效试次不足,因此被排除在脑电分析之外。所有受试者均为右利手,视力正常或矫正至正常,且无神经或精神疾病史,他们在实验前均签署了书面知情同意书。该研究获得了法兰克福歌德大学伦理委员会的批准,并根据《赫尔辛基宣言》进行。这些受试者是从当地大学招募的,每小时的报酬为15欧元。
视觉刺激
Presentation(V10.3,神经行为系统)用于刺激呈现和反应收集。所有刺激均使用Matlab(The Mathworks)生成。刺激以150×150像素矩阵的形式呈现在显示器屏幕中心,刷新率为60Hz,距离受试者的眼睛平面70cm处,视角为4.4°×4.4°,周围是灰色背景,在灰度[0,1]上的灰度级别为0.5。
EEG记录期间的任务
对于每个受试者,使用了160种不同的刺激。在每个试次开始时,先呈现一个十字注视点,随机持续时间为2~3s。然后,呈现一个Mooney图像8s。将此呈现视为试次的“第一阶段”。如果受试者能够识别图像中的物体,他们必须在这8s的间隔内尽快按下“Yes”按钮,从而终止试次。如果他们在8s内无法识别出物体,Mooney图像就会消失。然后,在1.5~2s的随机间隔后呈现一幅灰度图像作为线索/提示,此图片持续时间为4s。在50%的试次中,灰度图像与最后的Mooney图像一致,并作为后续识别的线索。在另外50%的试次中,它们是不一致的。本研究将这一灰度图像呈现视为试次的“第二阶段”。当灰度图像消失后,随机间隔2~3s,最后再次呈现一张Mooney图像。本研究将Mooney图像的重复呈现视为试次的“第三阶段”。在这个阶段,受试者必须尽可能快速准确地按下“Yes”或“No”按钮,以表明他们是否能够识别Mooney图像中的物体。响应需要在“第三阶段”图像呈现时间内完成(即最长8s;这个时间范围足以让受试者完成响应)。如果受试者在匹配试次中回答“Yes”,则将此视为他们经历了尤里卡效应的证据,即突然识别出他们在“第一阶段”无法识别的模式。图1说明了任务过程。试次顺序和实验条件的顺序是随机的。实验共分为4个block。每个block之间有2~3min的休息时间。在实验之前,使用不同的一组图像进行训练,以便让每个受试者进行练习。
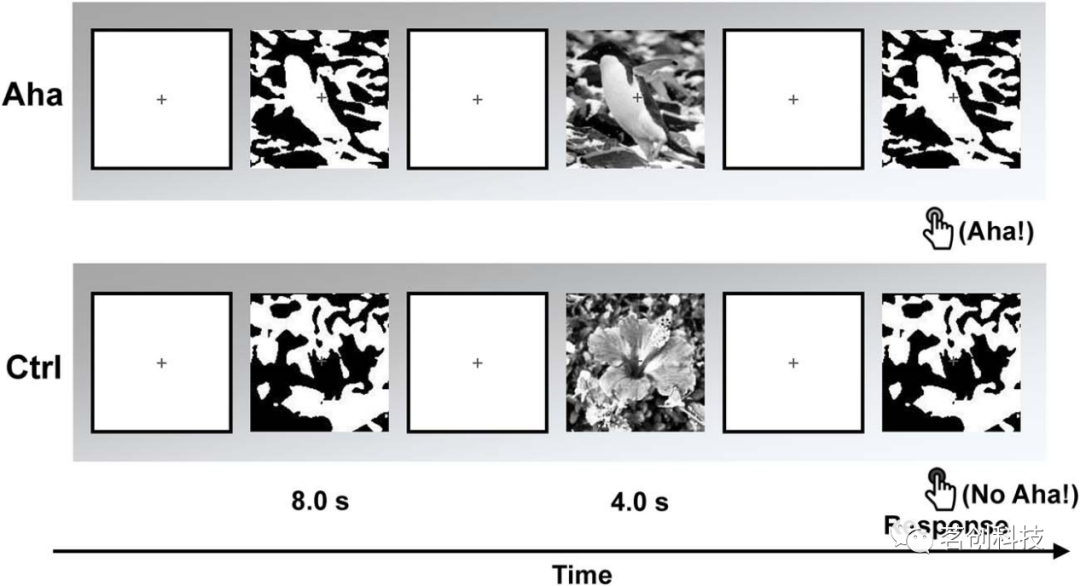
图1.EEG记录期间的范式。上行显示了尤里卡(Aha)试次的示例,该试次由Mooney图像及其一致的灰度图像组成。下行显示了对照(Ctrl)试次的示例,该试次由Mooney图像和不一致的灰度图像组成。
数据采集
EEG使用具有128个通道的HydroCel Geodesic Sensor Net 130(Electrical Geodesics,Inc.)进行记录,参考电极位于Cz。电极按照指示放置在头部上方。电阻保持<50kΩ。数据以1000Hz的频率采样,并以数字化方式保存在Mac系统中用于离线分析。
行为分析
本研究将试次分为两组:尤里卡(Aha)组和对照(Ctrl)组。Aha组由(线索一致)试次和“第三阶段”的“Yes”响应组成。Ctrl组由(线索不一致)试次和“第三阶段”的“No”响应组成。不符合这些要求的试次(9.5%的试次)从进一步的分析中排除。然后,本研究比较了Aha组和Ctrl组试次的反应时间(RTs)分布。
EEG数据预处理
在Net Station软件4.5.1(Electrical Geodesics,Inc.)中,对连续EEG信号进行高通滤波(0.3Hz)和陷波滤波(50Hz),然后将连续信号分割成一系列2-10s的epoch(取决于RTs)。每个试次段在“第三阶段”开始前1s开始,在“第三阶段”响应后1s结束。在接下来的步骤中,使用Matlab工具箱FieldTrip和本研究定制的脚本分析数据。通过目视检查、FieldTrip脚本和独立成分分析来补充半自动伪迹剔除,以检测电极漂移、眼动以及肌电图和心电图干扰。被拒绝的通道使用球面样条插值。为了避免刺激起始而不是尤里卡效应引起的神经活动污染,仅对RTs时间大于0.5s的试次进行分析。
频谱分析
为了研究尤里卡效应有关的时频活动,本研究采用汉宁锥度方法分析了整个电极空间中脑电信号的频谱变化,该方法具有5个周期的时间窗,频率平滑宽度随频率变化,步长为1Hz。分析时间窗为1s(对于刺激锁时的epoch,时间窗为-0.25~0.75s;对于响应锁时的epoch,时间窗为-0.75~0.25s)。对于基线校正,使用“第一阶段”刺激开始后的时间段,其长度与“第三阶段”中使用的分析窗长相同,其持续时间由各自的RTs确定。综上所述,Aha和Ctrl之间的基线校正和比较可以用一个简单的公式来描述:Diff=(M3Aha-M1Aha)-(M3 Ctrl-M1Ctrl),其中,Diff是Aha和Ctrl之间的差值;M3和M1分别为“第三阶段”和“第一阶段”的Mooney图像数据;M3-M1是指专门用于频谱分析的dB转换。减法去除了由刺激诱发反应产生的分量。最后,计算每个条件和参与者的总体平均值。Theta,alpha,beta和gamma频段的频率范围分别为:4-7、8-12、13-30和31-100Hz。
为了避免试次数量不相等可能带来的偏倚,在比较前,通过从试次数量较多的条件中随机丢弃试次(18.9%的试次),使Aha组和Ctrl组的试次数量相等。该校正也用于相干性和锁相值(PLV)分析。
相干性
大脑网络在解剖学和功能上都有定义。鉴于处理流之间的高度解剖连接,任何认知和执行任务都需要快速灵活地形成功能网络。功能连接的一个常用测量指标是相干性。它同时考虑了信号的相位和幅值分量,并提供了关于网络节点解剖和功能耦合的信息。因此,本研究计算了电极位置/通道之间在选定时间窗内特定频段的相干性。根据以下公式计算相干性:
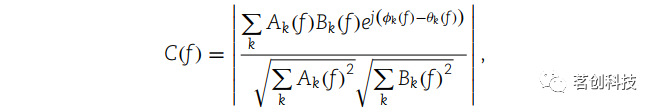
锁相值
由于相干值对振幅变化很敏感,因此本研究还计算了锁相值(PLV)。这些值反映了两个振荡信号之间的相位相关性,并作为不同神经元群同步的指标。区别PLV与相干性的关键因素是PLV不考虑振幅,它只测量相位分量。与相干性类似,PLV也有一个从0到1的实数值。1表示两个信号是严格锁相的,而0表示两个信号的相位关系是随机的。PLV可以表示为:

偏侧化指数
通过计算偏侧化指数(LIs)来获得偏侧化效应。通过评估左右半球之间的差异来计算LIs。使用以下公式进行计算:
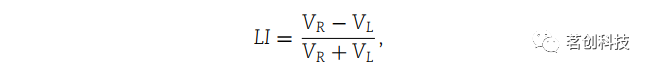
其中VR和VL是指左右半球或左右电极簇内的平均值。LI的正或负分别表示神经活动向右侧或左侧偏侧化。为了计算相干性和PLV的LI,只选用了一组电极。右电极:Fp2、F4、F8、C4、T4、P4、T6和O2;左电极:Fp1、F7、F3、T3、C3、T5、P3和O1;中线电极:Fz、CPz、Pz和Oz。右半球的相干性和PLV值是根据“右电极”内组合和“右电极”-“中线电极”组合计算的。同样,左半球的值是根据“左电极”内组合和“左电极”-“中线电极”组合计算的。根据10-20 EEG系统确定电极位置。
维数
为了评估网络动态复杂性的变化,本研究确定了记录的高维时间序列的分形维数。将EEG数据归一化为z分数,然后进行分形结构分析。对于大小为δ球体内的n个点(即这些点之间的距离<δ),

其中(xi,xj)是指数为i和j的点对(|xi-xj|<δ,i≠j),C(δ)为相关积分。维数D可由ln(C(δ))与ln(δ)的斜率来估计:

统计学
为了评估尤里卡效应与对照条件的试次之间差异的显著性,当样本为非正态分布时,进行Wilcoxon符号秩检验;或当样本服从正态分布时进行t检验(双尾)。采用基于聚类的非参数随机化检验来解决多重比较问题。对于每个配对样本进行独立样本t检验。选取P<0.05的所有样本,并根据时空邻接性进行聚类分析。计算聚类t值的总和。然后在Aha和Ctrl条件下随机分配这些样本,并分析聚类t值。此步骤重复1000次。然后得到一个新的聚类t值的分布。在此分布的基础上,对原始数据的t值进行评估。本研究使用0.05的显著性水平获得显著的数据簇(例如:包括预定义频率的电极和时间窗)。同样,基于聚类的非参数随机化检验也用于校正Wilcoxon符号秩检验的z值。
结果
本研究调查了尤里卡效应期间EEG记录的大脑活动的电生理特征。本节将在后续段落中描述行为结果、不同频段的功率变化、alpha和theta频段中相干性和PLV的变化,以及分形维数的变化。
行为数据
83.9%的试次持续到“第三阶段”,其中90.5%的试次反应正确。正确意味着对一致试次的响应为“Yes”或对不一致试次的响应为“No”。这一结果表明,受试者确实经历了尤里卡效应,并且Aha和Ctrl条件之间的比较是有效的。图2总结了尤里卡(Aha)和对照(Ctrl)试次的RTs。Aha试次中的RT明显快于Ctrl试次中的RT。RTs的平滑分布表明,Aha RTs分布的峰值在0.90s,而Ctrl RTs分布的峰值在1.50s。Aha RTs分布比Ctrl RTs分布更窄。Aha试次RTs的中位数为1.13s,Ctrl试次RTs的中位数为2.00s。在92.9%的Aha试次和99.3%的Ctrl试次中,RTs大于0.50s。这些试次被保留以供进一步分析。
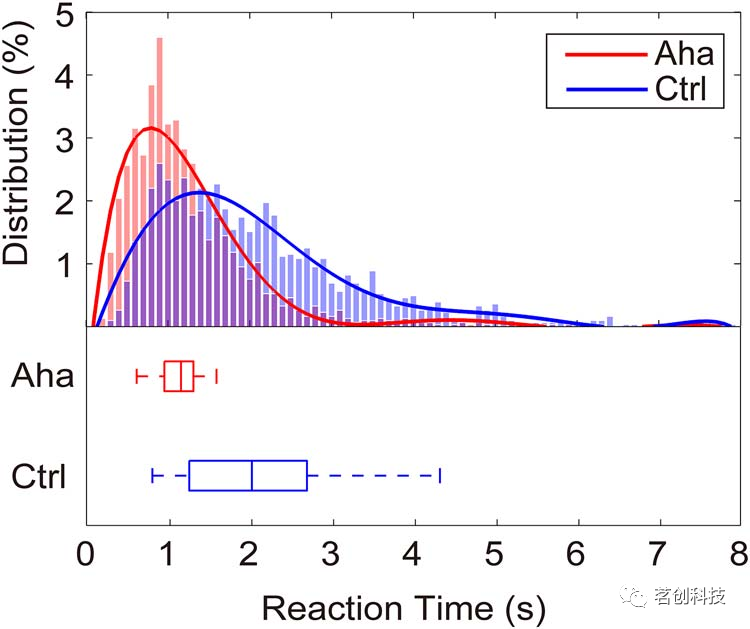
图2.RTs的分布。
频谱变化
Alpha功率
鉴于与尤里卡效应相关的许多功能和过程,如注意、记忆和自上而下的调制,都涉及alpha频段的活动,本研究探讨了alpha功率的变化是否与尤里卡效应有关。在刺激锁定时段,Aha和Ctrl条件下的alpha频段(8-12Hz)功率无显著差异。在反应锁定时段,Aha试次的alpha功率低于Ctrl试次。如图3所示,本研究发现了两个alpha功率降低的团簇(从基于聚类的非参数随机化检验得到)(P<0.05)。一个在额叶区域(主要在左侧额区),从反应前150ms (-150~0ms)出现。在该团簇中,alpha功率在反应之前显著降低。第二个团簇位于枕叶区域,从反应前335ms(-335~0ms)出现。有趣的是,随着时间的推移,这一团簇表现出从左半球到右半球的逐渐移动。在−220ms之前,该团簇主要位于左侧枕区;−220ms后,移至右侧枕区和右顶上区。简而言之,本研究观察到alpha功率下降与尤里卡效应有关。

图3.对诱发尤里卡效应的刺激做出反应之前alpha功率的变化。
Beta功率
尤里卡效应涉及许多认知过程,其中两个与beta功率的变化有关——记忆检索和认知状态的转换。因此,本研究考察了beta功率变化是否与尤里卡效应有关。在刺激锁定时段,尤里卡效应与刺激开始后240-360ms之间(相对于Ctrl)的beta功率(13-30Hz)下降有关(P<0.05),如图4所示。这种减少在靠近中线的右侧顶叶皮层和中央回区域最为明显。在反应锁定时段,基于聚类的非参数随机化检验显示,在反应前−380~−285ms的时间窗内,右侧顶叶和右侧枕叶皮层的beta功率有较为明显的下降趋势,但未达显著性水平(P= 0.0789)(图5)。总的来说,beta振荡功率的下降与尤里卡效应有关。
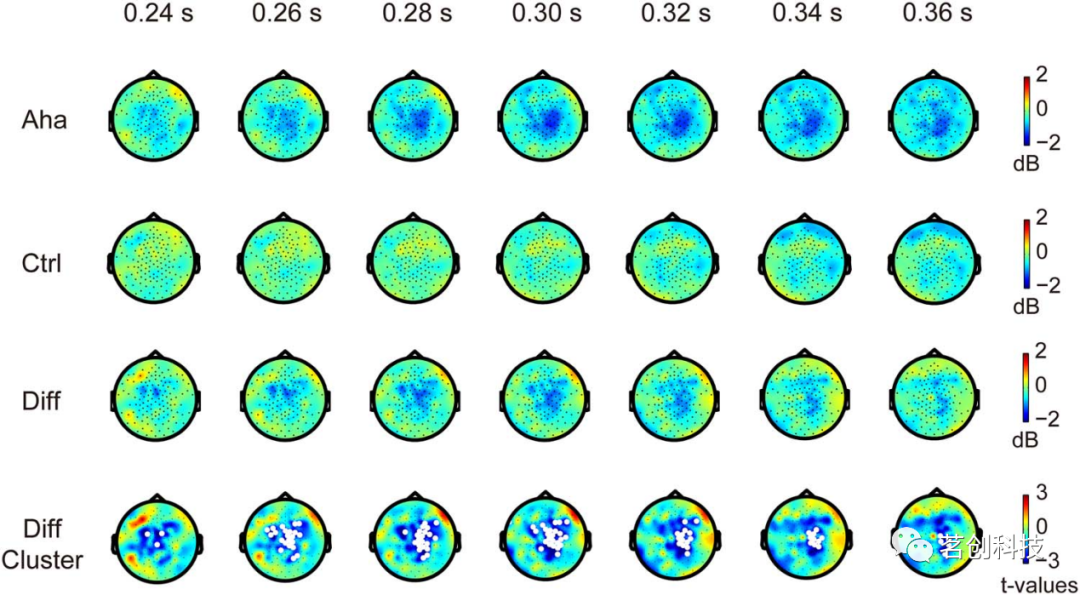
图4.刺激开始后beta功率的变化。

图5.对诱发尤里卡效应的刺激做出反应之前beta功率的变化。
Theta功率和gamma功率
Theta频段振荡被认为与各种记忆过程有关,而gamma频段振荡则与感觉证据和情境预测的整合有关。本研究考察了在尤里卡经历期间theta和gamma振荡的功率是否发生了变化。然而,这些频段的功率分析显示尤里卡和控制条件之间没有显著差异。
右半球的alpha和theta相干性增加
Alpha相干性
为了分析低频范围内通道之间的同步,本研究计算了从刺激开始(从0到500 ms,“刺激锁时”)时500ms窗口内的相干性。该分析结果显示,对于Aha条件,alpha频段的相干性增加,并具有右半球偏侧化(图6A)。为了量化这种偏侧化,本研究计算了通道对的alpha相干性的LI。这证实了在刺激后345至450ms的时间窗内,与Ctrl条件相比,Aha的右半球alpha相干性显著增加(P<0.05)。由于各试次的RTs不同,本研究还将分析窗口与响应对齐(从-500ms到反应开始,“反应锁时”)。右半球偏侧化alpha相干性的增加也出现在该反应段中,并且在-355~-275ms的时间窗内显著增加(P<0.05)(图6B)。
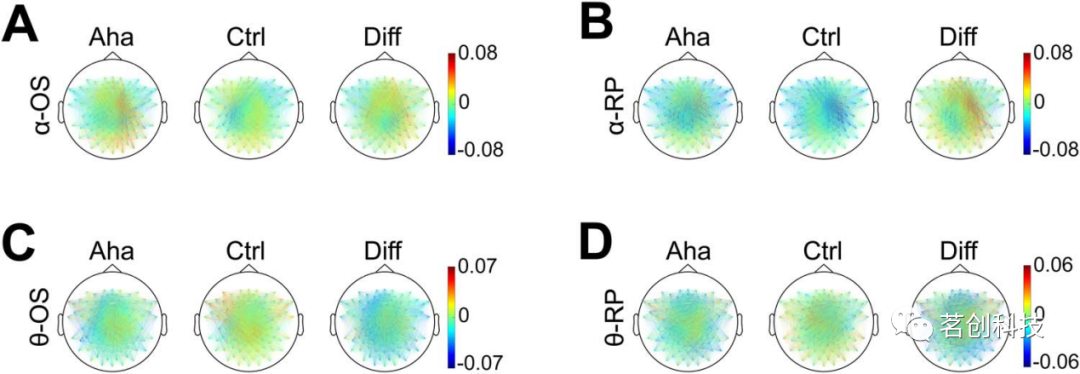
图6.alpha(A,B)和theta(C,D)相干性的地形分布。
Theta相干性
Theta相干性分析也显示,Aha条件在刺激后0-500ms内的右半球偏侧化增加(图6C)。LI分析表明,这种右侧化在两个时间窗内是显著的:125~260ms(P<0.05)和375~395ms(P<0.05)。对于反应锁定时段,只有右半球相干性呈增加趋势,但这种半球间差异不显著(图6D)。Theta相干性的iCOH分析表明,从刺激后180~365ms,与Aha相关的右侧化显著增加(P<0.05)。对于反应锁定时段,未观察到半球间的差异。
右半球alpha和theta的锁相增加
Alpha PLV
为了评估与尤里卡效应相关的锁相变化,本研究分析了刺激开始后0~500ms的alpha频段通道对的PLVs。在尤里卡效应期间,alpha频段的PLV也增加了,这种增加的地形(图7A)类似于alpha相干性的增加。在刺激后385~445ms,Aha右半球的PLV(相比于Ctrl条件)显著增加(P<0.05)。PLV在反应锁定时段也明显增强,同样右侧的增强程度高于左侧,但这种半球间差异未达到显著性(图7B)。iPLV的LI分析显示,刺激后315-425m和反应前500-395ms,右半球的iPLV明显增加(P<0.05)。

图7.Alpha和theta PLV的地形分布。
Theta PLV
刺激锁定时段的PLV分析再次显示,Aha试次中的右侧theta同步性增加(图7C)。LI分析表明,这种效应在375-420ms的时间窗内显著(P<0.05)。对于反应锁定时段,也存在右侧化增加的趋势,但LI指数未达到显著性水平(图7D)。Theta频段的iPLV证实,与Ctrl相比,Aha在刺激后370~405ms的右侧化显著(P<0.05)。在反应锁定时段没有发现半球间差异。
分形维数的变化
大脑可以被认为是一个表现出非线性动力学的复杂系统。因此,频谱分析可能无法捕捉到与认知状态切换相关的大脑活动的复杂性。在这里,本研究还采用了一种用于评估复杂动态系统状态的度量方法——分形维数。本研究分析了10个预定义的电极簇在刺激开始后0~500ms时间窗中的维数。在尤里卡试次中,可以观察到右侧中央和左侧顶叶区域的维数显著减少,右侧枕叶区域的维数显著增加(图8A)。此外,LI分析显示,右半球颞区降维更为明显(ln(δ)从1.47至1.65,P<0.05)(图8B)。与反应相关的活动模式也得到了类似的结果。在从−500ms到反应的时间窗中,右侧颞区的维数显著减小,左侧额叶区域的维数显著增加(图9A)。LI分析证实了颞区降维(ln(δ)从0.94至1.26,P<0.05)的右侧偏侧化(图9B)。

图8.刺激开始后500ms内活动向量维数的变化。
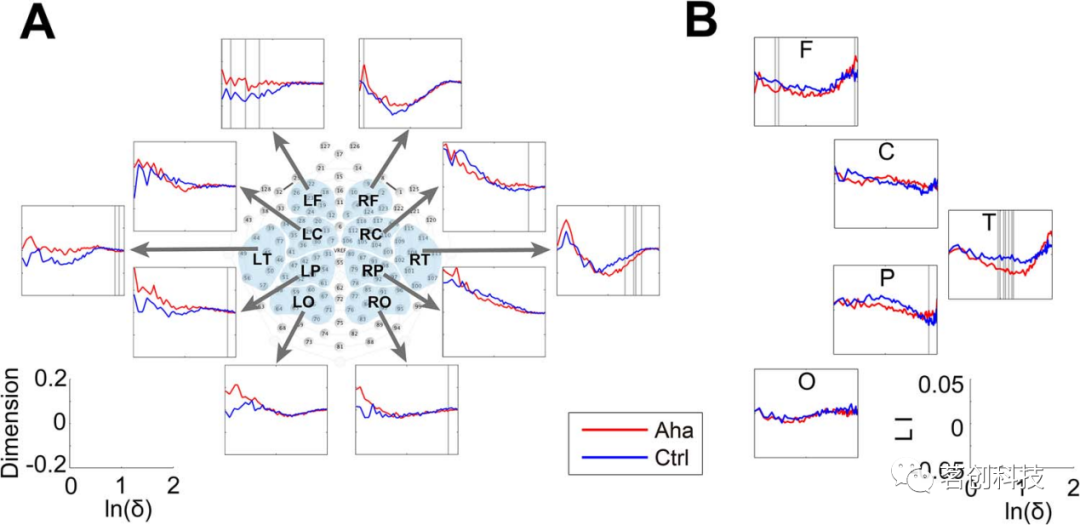
图9.对诱发尤里卡效应的刺激做出反应前500ms内活动向量维数的变化。
综上所述,在刺激锁定和反应锁定的时段,尤里卡效应与右侧颞叶皮层分形维数的减少有关。
结论
本研究的数据表明,尤里卡效应主要涉及右利手被试的右半球。本研究将alpha和theta频段中相干性的增加解释为形成广泛分布的皮层区域网络的指标,这些网络涉及将感觉证据与工作记忆中存储的信息进行比较。这一发现表明,增强的相干性与动态状态维数的降低有关,但与各自频段的功率增加无关,因为尤里卡的神经元关联经历了一个大网络向动态状态收敛的过程,其特征是维数减少、自由度降低,因此模糊性降低。
原文:Dynamic signatures of the Eureka effect: an EEG study.
https://doi.org/10.1093/cercor/bhad150