程序性细胞死亡 (Programmed cell death, PCD) 是多细胞生物中,由基因调控的细胞自杀过程,对多细胞生物的发育、体内稳态和完整性至关重要。PCD 的研究涉及多个领域,如免疫、神经系统发育、癌症、感染等。常见的 PCD 有细胞凋亡 (Apoptosis)、自噬 (Autophagy) 和焦亡 (Pyroptosis),以及近年发现的铁死亡 (Ferroptosis) 。
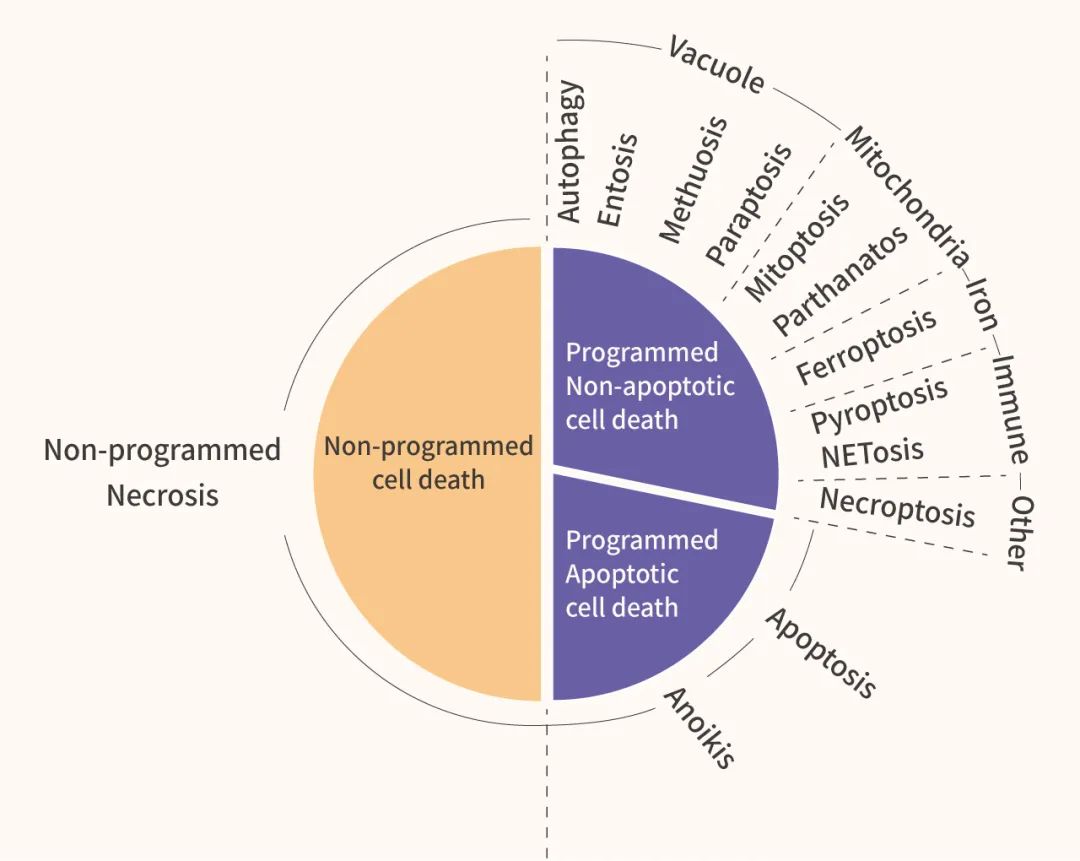
细胞焦亡 (Pyroptosis) 是一种促炎症的调节性坏死 (Regulated necrosis) 方式。细胞焦亡通常在细胞内病原体 (Intracellular pathogens) 感染时发生,例如单核细胞、巨噬细胞和树突状细胞被一系列微生物病原体 (如 Salmonella, Francisella, Legionella) 感染后,发生半胱氨酸蛋白酶 Caspase-1 依赖性细胞焦亡。此外,一些损伤相关的分子模式 (DAMPs) 也会引起非巨噬细胞的焦亡。
细胞焦亡主要有两种分子机制: 经典的 Caspase-1 依赖性和非 Caspase-1 依赖性途径,这两种机制的焦亡都会导致促炎症细胞因子 IL-1β 和 IL-18 释放 (图 2),扩大局部或全身的炎症。
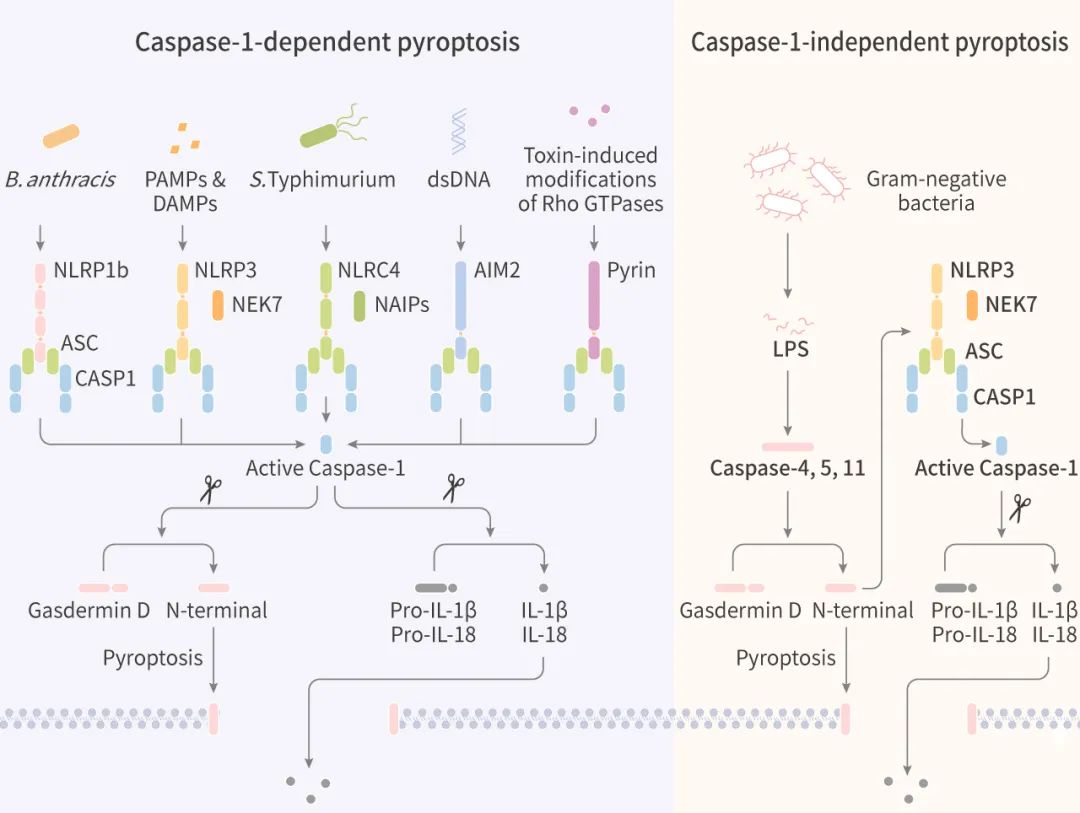
(左) Caspase-1 依赖性细胞焦亡;(右) 非 Caspase-1 依赖性细胞焦亡
在了解细胞焦亡机制之前,我们需要先认识两种重要组分: 炎症小体 & Gasdermin D。
炎症小体 (Inflammasome): 一般是由上游感受器蛋白,即模式识别受体 PRR,和衔接蛋白 ASC 以及下游的 Caspase-1 组成的复合物 (图 3),其中 PRR 能识别病原体入侵诱导的某些病原相关分子模式 PAMPs 和损伤相关分子模式 DAMPs。PRR 家族包括 Toll-样受体 (TLRs)、NOD-样受体和 AIM2 等成员,经典的炎症小体亚型的感受器包括 NOD-样受体 (NLRP1, NLRP3, NLRC4) 、AIM2 或热蛋白结构域 PYD,在识别 PAMPs 或 DAMPs 后活化 Caspase-1。
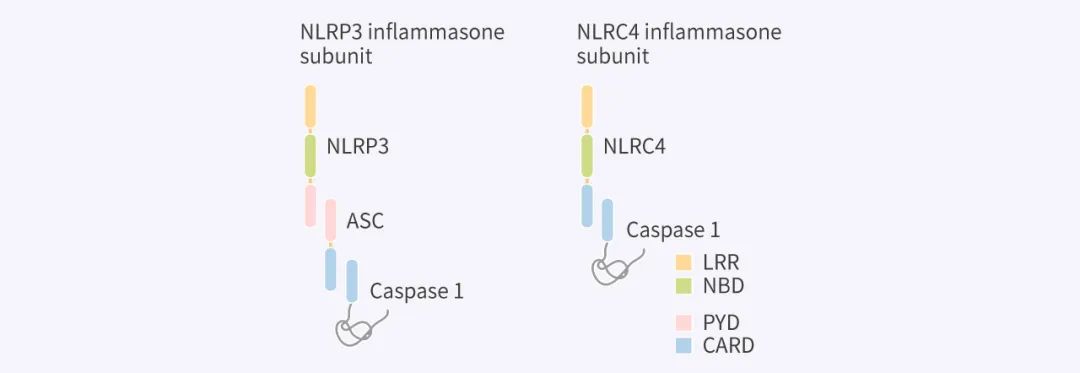
LRR: 富含亮氨酸重复序列;NBD: 核苷酸结合和寡聚结构域 ;PYD: 热蛋白结构域;CARD: Caspase 激活和募集域结构域 CARD
Gasdermin D (GSDMD): 焦亡中炎症 Caspase-1/4/5/11 的直接底物。GSDMD (53 kDa) 被 Caspase-1/4/5/11 裂解产生一个能启动焦亡的 N-末端片段 ( GSDMD-NT, 31 kDa) 和一个功能未知的 C-末端片段 (GSDMD-CT, 22 kDa)。GSDMD-NT 与细胞膜内小叶结合,并构成直径 10-33 nM 的孔。GSDMD-NT 也驱动 NLRP3-依赖性 Caspase-1 炎症小体的激活,这可能要求 GSDMD 诱导形成的孔隙并引起钾外排。
Caspase-1 依赖性细胞焦亡 (图 2. 左): 以 NLRP3 途径为例,某些 PAMPs 或 DAMPs 激活炎症小体感受器后,触发 ASC 和 Caspase-1 募集形成大分子复合物,在复合物中 Caspase-1 被激活 。激活的 Caspase-1 直接裂解 GSDMD 和前体细胞因子 pro-IL-1β 和 pro-IL-18,引发焦亡,并促使 IL-1β 和 IL-18 成熟。裂解的 GSDMD-NT 在细胞膜上形成孔,介导细胞质内容物 (Contents) 的释放。炎症小体感受器 NLRC4 与 NLRP3 不同之处在于 NLRC4 直接含有 CARD 结构域,可以直接与 Caspase-1 相互作用 (图 3)。非 Caspase-1 依赖性细胞焦亡 (图 2. 右): 人细胞中的 Caspase-4 或 Caspase-5 或小鼠细胞中的 Caspase-11 识别细胞溶质中的 LPS,这些炎性 Caspase 直接裂解 GSDMD 并引发焦亡。此外,GSDMD-NT 还能激活 NLRP3 炎症小体及 IL-1β 和 IL-18 的 Caspase-1 依赖性成熟。除了以上两种主要的细胞焦亡机制外,最近的研究还发现了一种新的焦亡途径: 特定的刺激导致 Caspase-3 的激活 (原本 Caspase-3 是凋亡的底物),Caspase-3 诱导 Gasdermin E (GSDME) 的裂解为 C-末端片段 (GSDME-CT) 和 N-端片段 (GSDME-NT)。GSDME-NT 也参与了膜孔的形成,导致细胞焦亡。
细胞凋亡 (Apoptosis): Caspases 依赖性的经典 PCD。凋亡细胞受控地分裂成凋亡小体,随后被周围的细胞和吞噬细胞识别和吞噬。在凋亡和焦亡中, Caspases 都发挥了重要的作用。
自噬性细胞死亡/自噬 (Autophagy-dependent cell death/Autophagy): 自噬体和自噬溶酶体的分解代谢过程,是细胞应激的重要保护机制,也是维持稳态的高度保守过程。
铁死亡 (Ferroptosis): 近年来发现的一种铁依赖性的 PCD,区别于细胞凋亡、细胞坏死、细胞自噬。
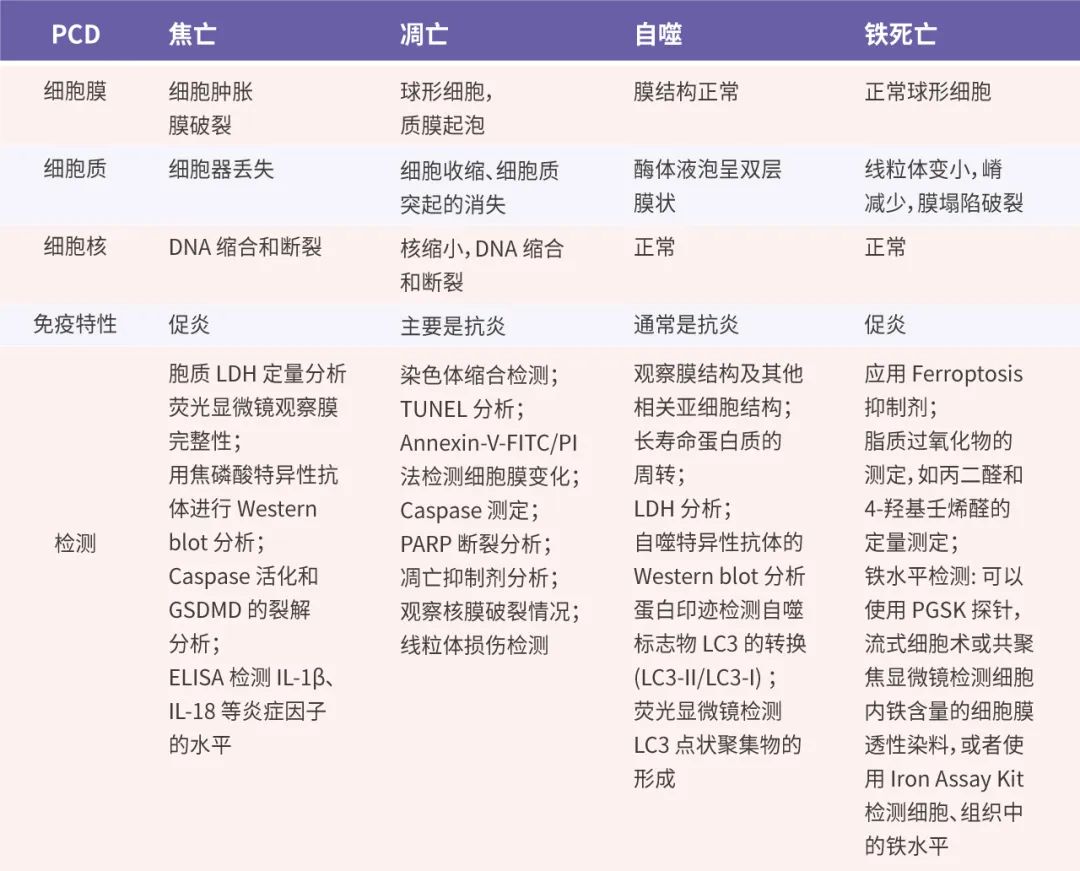
表 2. 几种 PCD 对比 (点击查看大图)[13]
细胞焦亡是机体重要的免疫反应,和感染性疾病、心血管疾病、神经系统相关疾病、肿瘤密切相关。细胞焦亡与感染: 病原体感染时,焦亡作用作为宿主防御机制,通过消除受损细胞,从而消除病原体,抵御感染,同时引发炎症反应。研究表明,多种细菌感染,包括弗氏志贺菌 (Shigella flexneri) 、沙门氏菌 (Salmonella)、单核细胞增生利斯特菌 (Listeria monocytogenes) 和嗜肺军团菌 (Legionella pneumophila) 都能导致细胞焦亡发生。细胞焦亡与心血管疾病: 近年来的研究表明,焦亡与动脉粥样硬化、心肌梗死、糖尿病性心肌病、再灌注损伤和心肌炎等心血管疾病的发病机制有关。例如,在早期的动脉粥样化形成中,内皮细胞的焦亡可能促进单核细胞招募进入内膜以及血管炎症;在晚期动脉粥样硬化病变中,巨噬细胞的焦亡可能促使坏死核心的形成和血小板不稳定。还有研究表明,心肌梗死期间心肌细胞的损伤与焦亡有关。细胞焦亡与神经系统疾病: 焦亡与中枢神经系统疾病如脑血管病、阿尔茨海默病和癫痫有关。例如阿尔茨海默病,其主要病理改变是细胞外神经炎斑块中 β-淀粉样蛋白 (Aβ) 的沉积干扰神经膜功能,导致钾离子从神经元中流出,激活 NLRP1,引发焦亡。
细胞焦亡与代谢性疾病: NLRP3 炎症小体在焦亡中起着重要作用,而 NLRP3 的激活可能与糖尿病的发生,发展及其相关并发症有关。
细胞焦亡与肿瘤: 近年来有研究发现,焦亡作用可影响肿瘤的增殖、侵袭和转移,并受到一些非编码 RNA 等分子的调控。
缩写:DAMPs: Danger-associated molecular patterns
PRR: Contain pattern-recognition receptors
ASC: Apoptosis-associated speck-like protein containing a CARD
PAMPs: Pathogen-associated molecular patterns
NLR: Nod-like receptor/Nucleotide-binding oligomerization domain-like receptors,CARD: N-terminal caspase recruitment domain
PYD: Pyrin domainLRR: leucine-rich repeat
NBD: Nucleotide-binding and oligomerization domain
AIM2: absent in melanoma 2
| 相关产品 | 作用 |
| 细胞焦亡化合物库 Pyroptosis Compound Library | 收录了 400+ 种细胞焦亡相关产品,主要靶向焦亡信号通路中主要靶点,可以用于细胞焦亡信号通路及相关疾病的研究 |
| 细胞凋亡化合物库 Apoptosis Compound Library | 收录了 1000+ 种凋亡相关产品,主要靶向凋亡信号通路种主要靶点,可以用于细胞凋亡信号通路及相关疾病的研究描述 |
| 自噬化合物库 Autophagy Compound Library | 提供 900+ 种自噬信号通路相关的产品,是研究自噬相关调控及疾病的有用工具 |
| 铁死亡化合物库 Ferroptosis Compound Library | 收集了 400+ 铁蛋白信号通路相关化合物,具有铁蛋白诱导或抑制活性。是研究铁作用机理的有用工具 |
| CCK-8 试剂盒 Cell Counting Kit-8 | 基于 WST-8 的广泛应用于细胞活性和细胞毒性检测的快速、高灵敏度试剂盒。 |
| JC-1 试剂盒 JC-1 Mitochondrial Membrane Potential Assay Kit | 以 JC-1 为荧光探针,快速灵敏地检测组织、细胞或纯化的线粒体跨膜电势差的试剂盒,可以用于早期的细胞凋亡检测 |
| 7-Aminoactinomycin | 荧光 DNA 染色剂 |
| Propidium Iodide | 用于细胞染色的红色荧光染料 |
| Erastin | 铁死亡诱导剂 |
参考文献
1. Mirosław Godlewski , et al. Programmed Cell Death - Strategy for Maintenance Cellular Organisms Homeostasis. Postepy Hig Med Dosw (Online). 2016 Dec 20;70(0):1229-1244.
2. Ge Yan, et al. Multiple cell death modalities and their key features (Review). WORLD ACADEMY OF SCIENCES JOURNAL. 2: 39-48, 2020.
3. Si Ming Man, et al. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases.Immunol Rev. 2017 May;277(1):61-75.
4. Jianjin Shi, et al. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem Sci. 2017 Apr;42(4):245-254.
5. Douglas R Green. The Coming Decade of Cell Death Research: Five Riddles. Cell. 2019 May 16;177(5):1094-1107.
6. Yuan Fang, et al. Pyroptosis: A New Frontier in Cancer. Biomed Pharmacother. 2020 Jan;121:109595.
7. Susan L Fink, et al. Apoptosis, Pyroptosis, and Necrosis: Mechanistic Description of Dead and Dying Eukaryotic Cells. Infect Immun. 2005 Apr;73(4):1907-16.Linde Duprez, et al. Major Cell Death Pathways at a Glance. Microbes Infect. 2009 Nov;11(13):1050-62.
8. Yuan Fang, et al. Pyroptosis: A new frontier in cancer. Biomed Pharmacother. 2020 Jan;121:109595.
9. Yaron Fuchs, et al. Live to Die Another Way: Modes of Programmed Cell Death and the Signals Emanating From Dying Cells. Nat Rev Mol Cell Biol. 2015 Jun;16(6):329-44.
10. Tessa Bergsbaken, et al. Pyroptosis: host cell death and inflammation. Nat Rev Microbiol. 2009 Feb;7(2):99-109.
11. Sushmita Jha, et al. Inflammasome-Associated Nucleotide-Binding Domain, Leucine-Rich Repeat Proteins and Inflammatory Diseases. J Immunol. 2009 Dec 15;183(12):7623-9.
12. J C Ameisen. On the Origin, Evolution, and Nature of Programmed Cell Death: A Timeline of Four Billion Years. Cell Death Differ. 2002 Apr;9(4):367-93.
13. Gladys O Latunde-Dada. Ferroptosis: Role of Lipid Peroxidation, Iron and Ferritinophagy. Biochim Biophys Acta Gen Subj. 2017 Aug;1861(8):1893-1900