今天给同学们分享一篇非肿瘤+线粒体+实验的生信文章“Role of mitochondrial metabolic disorder and immune infiltration in diabetic cardiomyopathy: new insights from bioinformatics analysis”,这篇文章于2023年2月1日发表在J Transl Med期刊上,影响因子为8.44。

糖尿病性心肌病(DCM)是糖尿病常见的心血管并发症之一,也是糖尿病患者死亡的主要原因。线粒体代谢和免疫炎症是扩张型心肌病发病机制的关键,但它们在扩张型心肌病中的相互作用仍然是一个悬而未决的问题。本研究利用生物信息学探讨了线粒体代谢和免疫微环境的各自作用及其在扩张型心肌病中的相互作用。
1. DCM 和功能富集分析中的DEG
整体数据筛选策略的流程图如图1所示。获取3个DCM相关GEO数据集GSE4745、GSE5606和GSE6880进行分析。差异分析显示,与正常样品相比,GSE4745数据集中有293个DEGs,其中DCM样品中有149个基因上调,144个基因下调;GSE5606数据集中的544个DEGs,包括269个上调基因和275个下调基因;以及GSE6880数据集中的463个DEG,包括262个上调基因和201个下调基因。DEG被可视化为火山图和热图(图2a-f)。
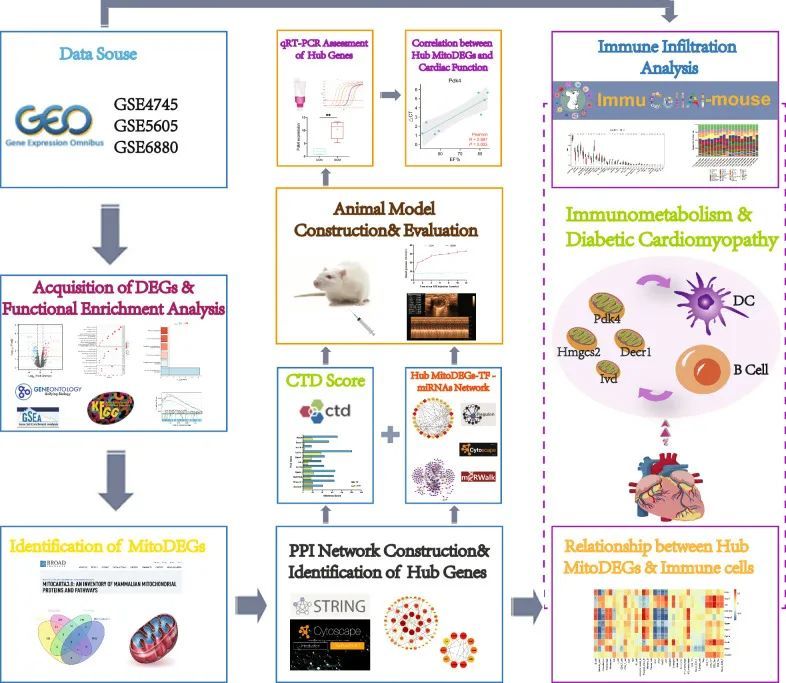
图1 流程图
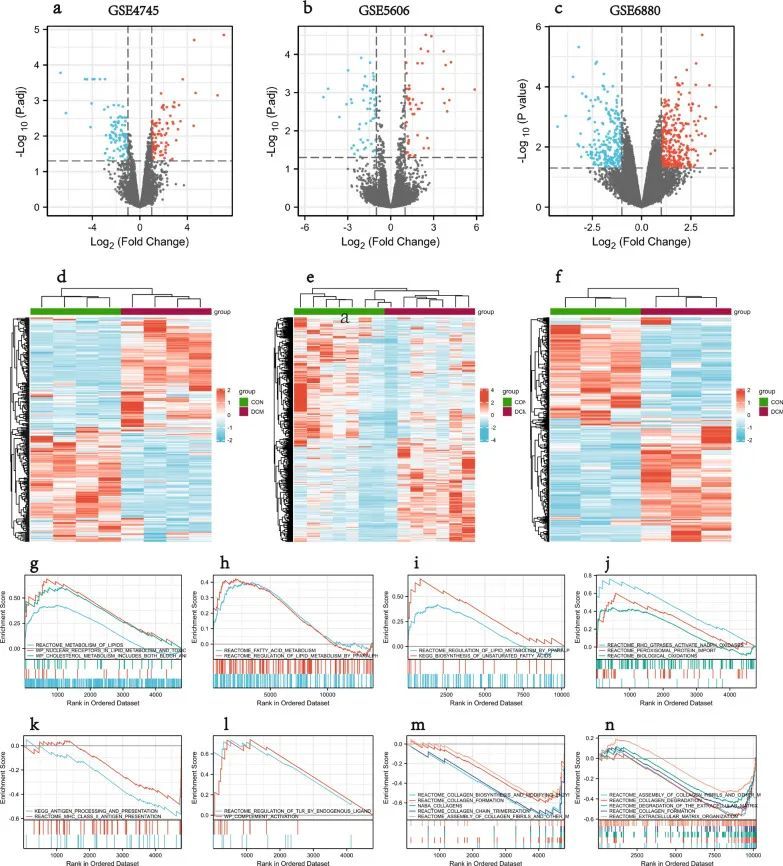
图2 DCM中的DEG和GSEA分析结果
GSEA显示,3个数据集的DEGs主要参与脂质和脂肪酸代谢以及免疫相关的途径,包括脂质代谢、PPARα调节脂质代谢、脂肪酸代谢、不饱和脂肪酸的生物合成、抗原加工和呈递、MHC II.类抗原呈递、内源配体对TLR的调控, 补体活化(图2g–n)。此外,它还显示出参与胶原蛋白合成、胶原纤维组装和氧化应激的途径的富集。
通过GO和KEGG途径分析进一步处理DEG以进行功能富集。GO术语最丰富的分为生物过程(BP)、细胞成分(CC)和分子功能(MF),主要包括线粒体功能和成分、能量代谢、炎症免疫、缺氧和氧化还原反应、胶原蛋白合成和胰岛素敏感性等。(图 3a–f)。DEGs最丰富的KEGG通路以线粒体代谢和功能、缺氧和氧化还原反应、物质生成和免疫等途径为主。(图 3g–l)。
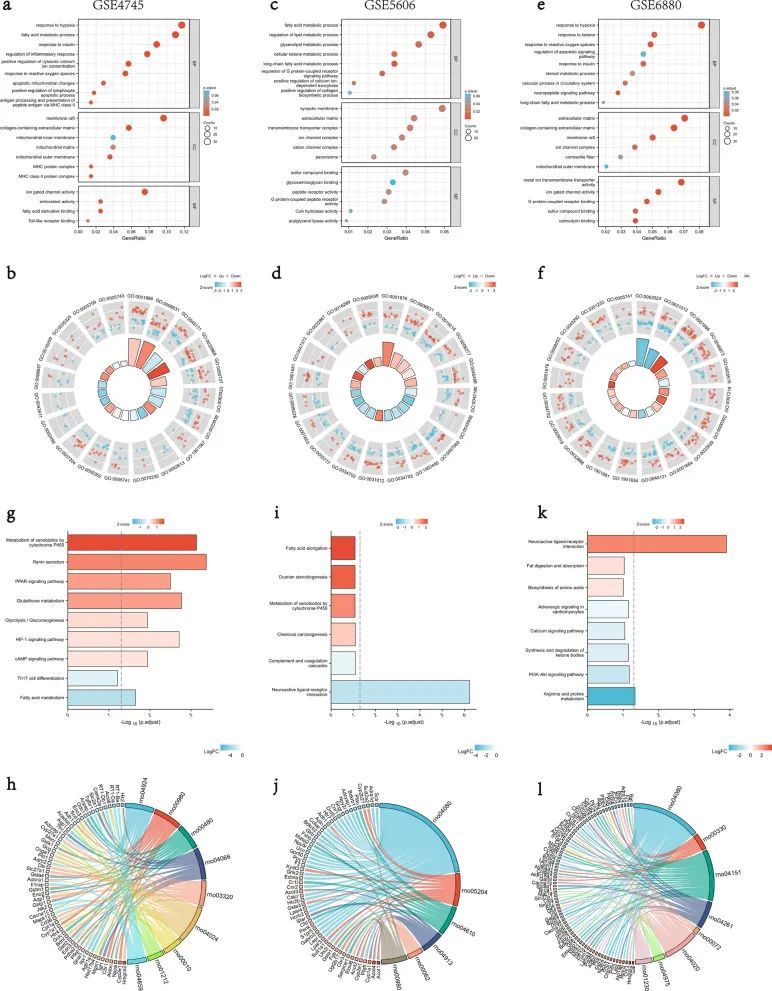
图3 来自 GSE4745、GSE5606 和 GSE6880 的 DEG 的 GO 和 KEGG 富集分析
2. DCM 中的 MitoDEG
从MitoCarta3.0数据库中检索线粒体相关基因,选择与3个数据集中的DEGs重叠的基因作为MitoDEGs。GSE4745数据集中总共有32个MitoDEC(15个上调和17个下调)(图4c),GSE5606数据集中有34个MitoDEP(18个上调和16个下调)(图4d),GSE6880数据集中有25个MitoDEG(14个上调和11个下调)(图4e)。合并每个数据集的MitoDEGs,得到67个交集的MitoDEGs,其中DCM样本中35个基因上调,32个基因下调。
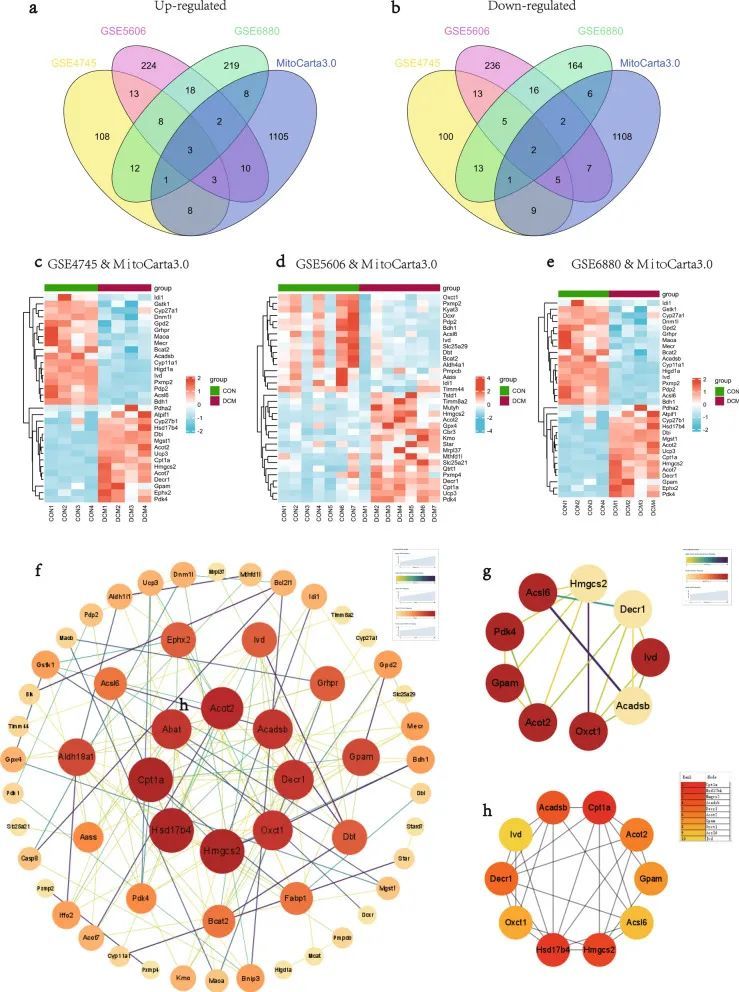
图4 DCM中的MitoDEGs、PPI 网络分析和关键 MitoDEG 识别
3. PPI 网络分析和关键 MitoDEGs 识别
使用STRING数据库分析67个MitoDEG的PPI,并使用Cytoscape可视化为网络(图4f)。使用插件MCODE鉴定重要模块。由9个节点和17个边缘组成的模块被鉴定为显著的,参与该模块的基因为Acsl6,Acadsb,Decr1,Ivd,Oxct1,Gpam,Pdk4,Hmgcs2和Acot2(图4g)。利用Cytoscape的MCC算法,从PPI网络中鉴定出10个候选关键基因,包括Cpt1a、Hsd17b4、Hmgcs2、Acadsb、Decr1、Acot2、Gpam、Oxct1、Acsl6和Ivd(图4h)。结合结果,最终获得了11个关键MitoDEG,包括Acadsb,Hmgcs2,Hsd17b4,Gpam,Acot2,Ivd,Decr1,Cpt1a,Acsl6,Oxct1和Pdk4。
4. 关键 MitoDEG 与 DCM/HF 之间的关系
利用CTD数据库预测关键MitoDEG与DCM/HF之间的关系。通过分析,Cpt1a、Gpam、Hmgcs2和Acadsb与DCM的相关性最高(图5a),而Cpt1a、Pdk4、Gpam和Hmgcs2与HF的相关性最高(图5b)。
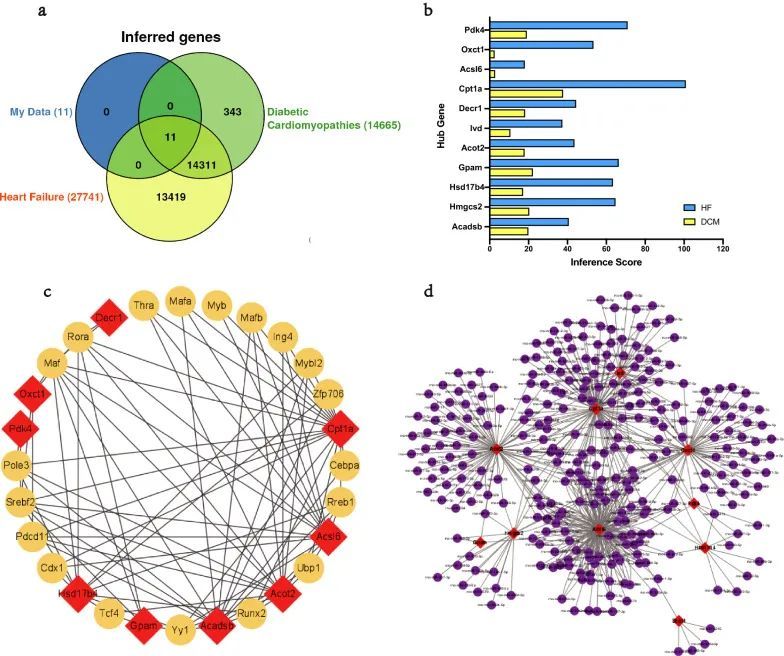
图5 关键 MitoDEG 与 DCM/HF 之间的关系
5. 关键 MitoDEGs-TFs-miRNAs 监管网络
通过预测相关的TFs和miRNAs,探索了中心MitoDEGs的上游调控。使用Cytoscape的插件iRegulon预测了关键 MitoDEG的TF,并构建了由19个TF(Rora,Maf,Ing4,Srebf2,Mafb,Zfp706,Pole3,Rreb1,Mybl2,Myb,Tcf4,Mafa,Cebpa,Thra,Pdcd11,Yy1,Runx2,Cdx1,Ubp1)组成的集线器MitoDEGs-TFs调控网络(图5c)。使用miRWalk 3.0预测了关键 MitoDEG的miRNA,并生成了涉及299个节点和569条边的MitoDEGs-miRNAs调控网络(图5d)。
6. DCM中的免疫细胞浸润
使用ImmuCellAI算法分析了36种免疫细胞类型的浸润,并在GSE5606和GSE6880数据集中比较了DCM和CON组。DCM组和CON组在9种免疫细胞类型的心肌浸润中存在显著差异。具体来说,DCM组中的B细胞,边缘区B和记忆B更丰富,而粒细胞,树突状细胞,MoDC,cDC1,pDC和cDC2在CON组中更丰富(图6a-c)。对DCM中浸润免疫细胞的进一步分析显示细胞之间存在多种相关性(图6d)。相关性程度由分数表示。CD4 T细胞与Naive CD4 T细胞(0.99)的协同效应最强,其次是CD4 T细胞和T辅助细胞(0.98)、CD8 Tcm和CD8 Tex(0.98)、Naive CD4 T和T辅助细胞(0.97)。相比之下,Naive CD8 T和B细胞之间的竞争效应最强(-0.72),其次是pDC和边缘区B(-0.69),Naive CD8 T和记忆B(-0.69)。
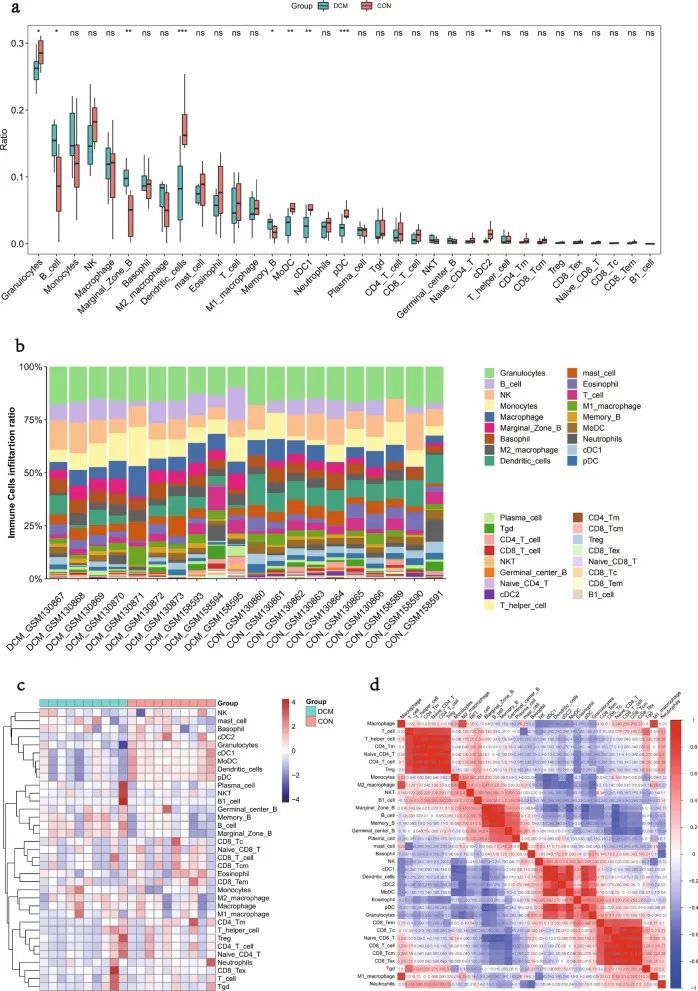
图6 DCM 和 CON 之间免疫细胞类型的浸润比较
7. MitoDEGs/关键 MitoDEGs与免疫细胞的关系
应用 Spearman 方法探索 MitoDEGs/关键 MitoDEGs 与免疫细胞之间的潜在关联。MitoDEGs(35 个上调和 32 个下调)与免疫细胞之间的正/负关联如图 7a、b 所示。在 11 个关键 MitoDEG 中,Pdk4 与 cDC2、MoDC 和 pDC 呈负相关;Oxct1 与 pDC 和 CD8 Tem 呈正相关;Ivd与CD8 Tem呈正相关;Hsd17b4 与边缘区 B 和 M2 巨噬细胞呈正相关,但与 cDC2、MoDC 和 pDC 呈负相关;Hmgcs2与边缘区B、M2巨噬细胞呈正相关,但与粒细胞和cDC2呈负相关;Gpam与树突状细胞、粒细胞、cDC1和MoDC呈负相关;Decr1与边缘区B和M2巨噬细胞呈正相关,而与粒细胞、cDC2和pDC呈负相关;Cpt1a与树突状细胞、cDC1、MoDC和pDC呈负相关;Acsl6 与 pDC、嗜酸性粒细胞和 CD8 Tem 呈正相关;Acot2与树突状细胞和cDC1呈负相关(图7c)。
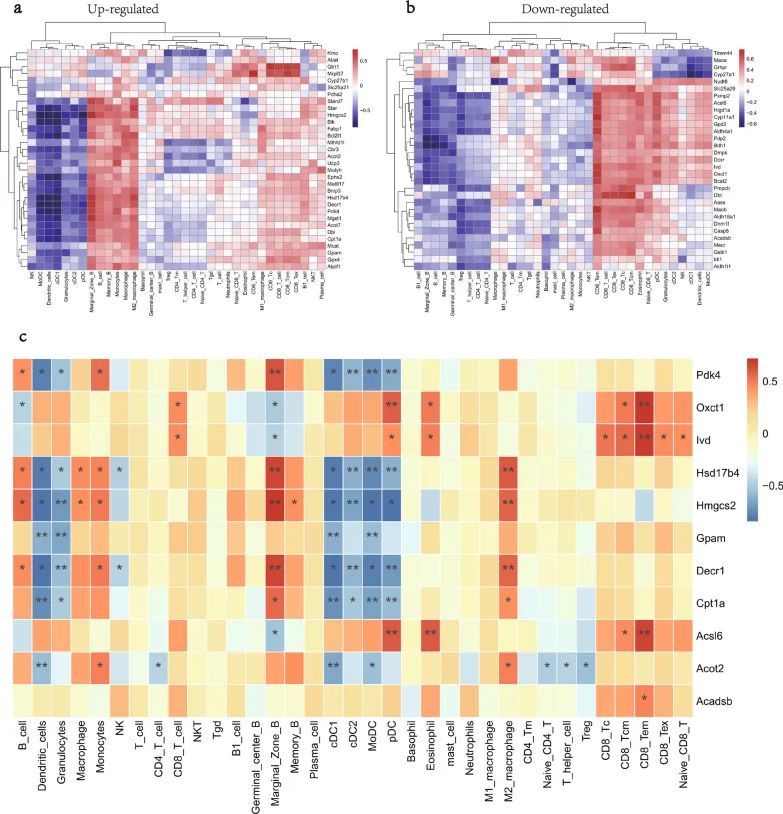
图7 MitoDEGs/关键 MitoDEGs与免疫细胞的关系
8. 扩张型心肌病大鼠的一般生物学和超声心动图特征
建模过程中,DCM组高脂日粮喂养大鼠体重显著高于CON组,且较STZ注射后2周起趋于下降,组织取样前明显低于CON组(图8a)。STZ诱导1周后,DCM组的血糖开始升高,并且在整个建模过程中水平始终高于CON组(图8b)。超声心动图显示,与CON组相比,DCM组的EF%和FS%显著降低(P < 0.05),但LVIDs显著升高(P < 0.05)。此外,两组之间的LVIDd略有变化(图8c-h)。此外,与CON组相比,DCM组的心脏重量(HW/BW)和心脏重量(HW/TL)显着增加(P < 0.05,图8i,j)。
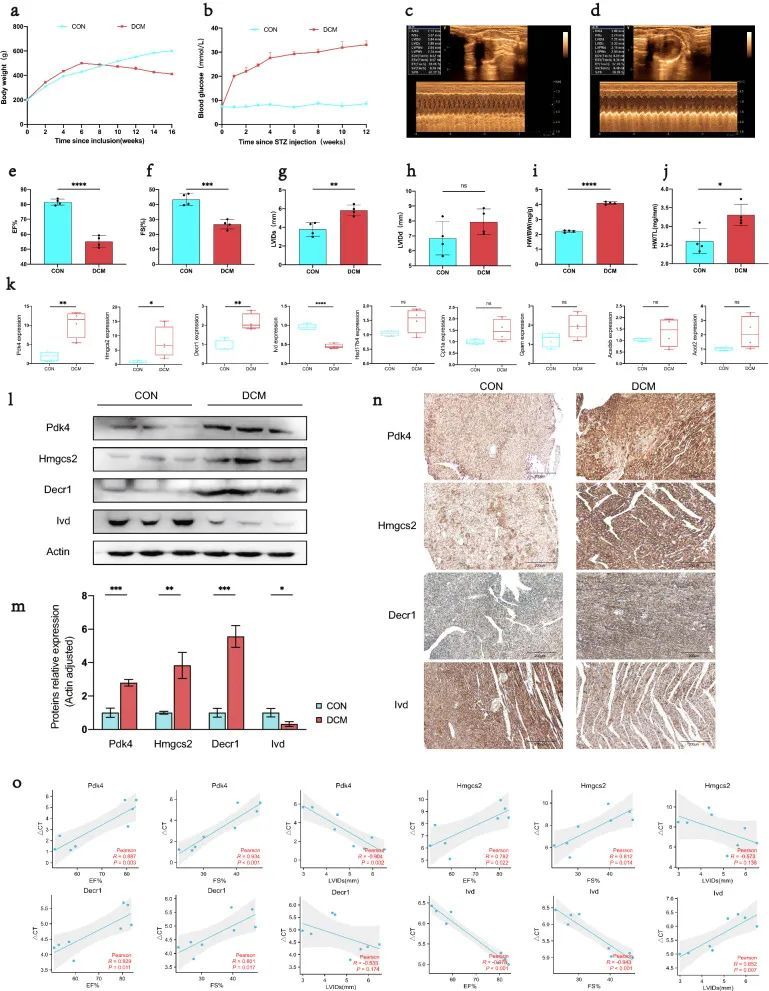
图8 确认DCM大鼠关键 MitoDEGs表达及其与心脏功能的关联
9. DCM 大鼠关键 MitoDEGs 表达的实验验证
9个关键MitoDEG(Acadsb,Acot2,Cpt1a,Decr1,Gpam,Hmgcs2,Hsd17b4,Ivd和Pdk4)的心室表达在qRT-PCR大鼠中得到验证。与CON组相比,Pdk4、Hmgcs2和Decr1在DCM组中的表达量显著增加(P < 0.05),而Ivd在DCM组中的表达量相反地表现出显著降低(P < 0.05)(图8k)。之后,通过蛋白质印迹和免疫组化进一步验证了DCM和CON组之间Pdk4、Hmgcs2、Decr1和Ivd的蛋白表达。结果表明,Pdk4、Hmgcs2、Decr1和Ivd的蛋白表达水平与mRNA的表达水平一致(P < 0.05)(图8l–n)。
10. 关键MitoDEG与心脏功能的关系
进一步分析了DCM和CON组之间表达明显差异的4个关键MitoDEG(Pdk4,Hmgcs2,Decr1和Ivd)与EF%,FS%和LVID的相关性。Pdk4的PCR循环次数与EF%呈极显著正相关(R = -0.904;P = 0.002)和满量程% (R = 0.934;P < 0.001),但与LVIDs呈极显著负相关(R = 0.852;P = 0.007);Hmgcs2的PCR循环次数与EF%呈极显著的正相关(R = 0.782;P = 0.022)和满量程%(R = 0.812;P = 0.014);Decr1的PCR循环次数与EF%呈极显著正相关(R = 0.829;P = 0.011)和EF%(R = 0.801;P = 0.017)。lvd的PCR循环数与EF%(R =⟩-0.978;P < 0.001)和FS%(R = -0.943;P < 0.001)呈极显着负相关,而与LVIDs呈极显着正相关(R = 0.852; P = 0.007) (图 8o)。总的来说,DCM 心肌组织中 Pdk4、Hmgcs2 和 Decr1 的上调表达以及 Ivd 的下调表达与心功能下降高度相关。
总结
综上所述,通过全面的生物信息学分析,确定了DCM和CON之间线粒体相关基因和免疫细胞浸润的差异。我们首次在DCM中发现了线粒体代谢和免疫浸润之间的串扰,筛选验证了4个关键基因,其中Pdk4、Hmgcs2和Decr1在DCM中表达量高,而Ivd表达率低。最重要的是,Pdk4、Hmgcs2和Decr1与边缘区B呈正相关,与直流电池呈负相关;IVD与CD8 Tem呈正相关。这些发现表明Pdk4,Hmgcs2,Decr1和Ivd是DCM中免疫代谢的共同调节分子。此外,边缘区B区、边缘区B区和CD8Tem的浸润差异在DCM的病理生理学中起着重要作用。