导读
本文提供了BOLD信号生理过程的总体概述(即生成生物物理模型),包括它们在生理信息动态因果模型(P-DCM)框架下的时间过程特征。BOLD信号主要由顺磁性脱氧血红蛋白的变化决定,而顺磁性脱氧血红蛋白的变化是氧代谢、脑血流量和脑血容量变化共同作用的结果。具体来说,本文讨论了所谓的BOLD信号“瞬态”的生理起源,包括初始过冲,稳态激活和刺激后下冲。在BOLD信号的生成模型中,错误的生理假设可能导致有关局部神经元活动和脑区之间有效连接的错误推断。此外,本文介绍了最近的层状BOLD信号模型,该模型将P-DCM扩展到皮层深度分辨率的BOLD信号,从而允许使用高分辨率fMRI数据来确定层状神经元的活动。
前言
典型功能神经成像实验的目标是绘制出受试者在执行任务、经历感官刺激或处于静息状态时的神经元激活模式。在MRI中,应用最广泛的对比是血氧水平依赖(BOLD)信号。它源自血管系统,因此是神经元活动的间接反映。BOLD信号通常使用一般线性模型(GLM)进行分析,用所谓的设计矩阵(即,与标准血流动力学反应函数(HRF)卷积的实验设计)回归每个体素的时间进程。这种方法产生了整个大脑BOLD活动的空间统计地图。
BOLD信号也用于推断功能连接和有效连接。功能连接被简单地定义为静息状态下体素时间进程的(瞬时)相关性。也就是说,在静息状态下,具有高度BOLD时间序列相关性的两个脑区被假定为在神经元水平上存在功能连接,并属于同一认知网络。因此,它隐含地假设在BOLD信号中观察到的所有变化都源自神经元,并且在功能连接的神经元中同步发生。
相比之下,有效连接被定义为一个大脑区域对另一个大脑区域产生的因果影响。最突出的有效连接方法是动态因果模型(DCM)。DCM利用局部生成生物物理模型描述了从神经元活动到BOLD信号的事件链,以及在神经元水平上将不同脑区之间因果关联的图形模型。在实验设计中,图形模型表示关于大脑如何处理某些任务的特定认知假设。DCM可以看作是GLM的扩展,其中包含了兴趣区域(ROIs)之间的交互项。因此,从生理和认知的角度来看,它比GLM更现实可行,因为刺激并不直接进入和影响体素的激活,而是通过平行和分层的途径,这可以用图形来表示。
BOLD反应的现象学
任务期间的BOLD信号与基线相比具有一些特征性的时间波动,也称为“瞬态”:它相对于神经元活动缓慢增加,并在4-10s后达到最大值。在刺激结束后,通常会观察到刺激后的下冲(见图1)。此外,还可能存在其他较少研究的瞬态:刺激开始和结束时的增加、初始过冲和BOLD负反应。在极少数情况下,会观察到BOLD信号的“初始下降”。刺激诱发的BOLD反应在受试者、脑区甚至相邻体素之间的振幅和时间进程上都有可能不同。
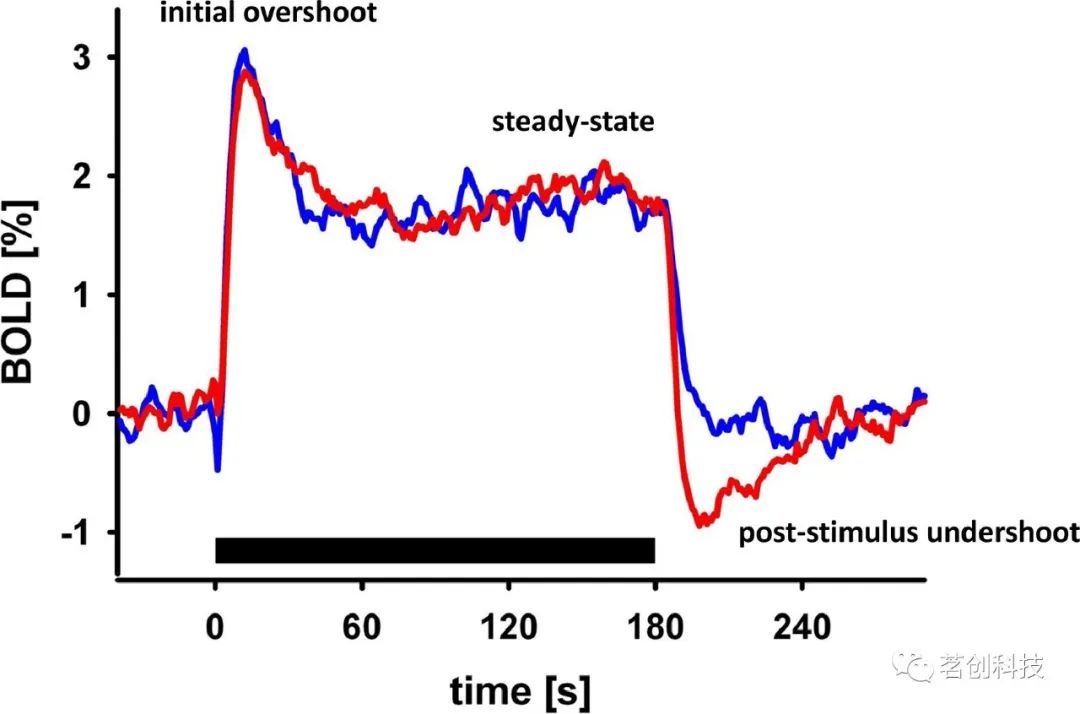
图1
BOLD信号是复杂的生理和物理过程的结果,取决于许多生理因素,例如基线血氧和容量,以及MRI采集参数(如回波时间、磁场强度和脉冲序列)。因此,这些BOLD信号瞬态在其生理起源方面是模糊的,这使得数据的解释更加困难。然而,这些经常被忽视的瞬态(例如,初始过冲和刺激后的下冲)可以提供关于神经元和血管反应复杂性的有趣见解,这些见解可用于研究脑生理学和统计脑图之外的信息加工。接下来,将讨论与有效连接建模最相关的BOLD信号瞬态的生理特征和起源。
刺激期间BOLD信号增加
由于脱氧血红蛋白(dOHb)具有顺磁性,因此血氧的变化会改变血液的磁化率,这可以通过T2和T2*加权MRI序列(分别为自旋回波和梯度回波fMRI)检测到。需要注意的是,顺磁性dOHb引起的血管磁化率变化会导致血管内和周围的质子移相。因此,尽管BOLD信号是由血管内变化引起的,但它同时具有血管内和血管外的MRI信号贡献。
神经元和星形胶质细胞的激活与氧化代谢(脑氧代谢率(CMRO2))的变化有关,氧化代谢增加了dOHb的含量(即血氧降低),并增加了磁化率。含有dOHb的血管的脑血容量(CBV)变化也会增加dOHb的含量。因此,CMRO2和CBV对产生的BOLD信号具有类似的影响。然而,随着脑血流量(CBF)的增加,从动脉流入的含氧血会过度补偿dOHb的增加。因此,CMRO2、CBV和CBF的联合作用导致dOHb整体降低,血氧增加(即BOLD信号)(图2)。
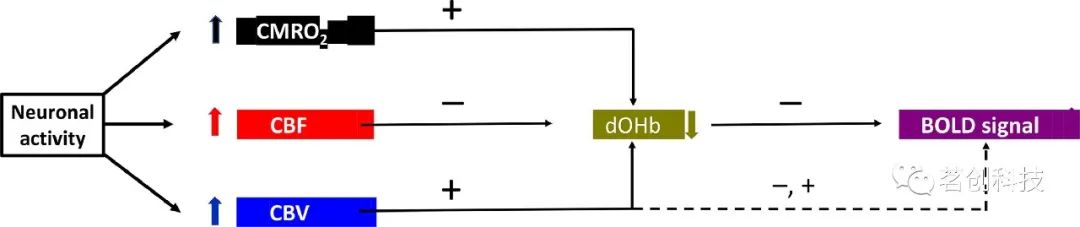
图2.导致BOLD信号的生理事件链。
重要的是,这些生理和物理变量的不同组合(例如,由于血管密度、神经血管耦合或磁场强度的差异)可以导致相同的BOLD信号稳态振幅。例如,balloon和Davis模型中BOLD信号方程的比例因子包含基线CBV0和磁场强度B0。因此,从高CBV0体素中获得的低B0的BOLD数据可以具有与从低CBV0中获得的高B0体素相同的BOLD信号灵敏度。
block设计的初始过冲
在许多fMRI数据集中,在信号达到一个新的稳态值之前,可以观察到持续刺激期间的初始过冲(见图1)。这种过冲通常被解释为神经元的起源。然而,如果静脉的CBV变化缓慢并在CBF已经达到稳态后继续增加,即使在没有神经适应的情况下,也会出现初始过冲。因此,在过冲后达到一个新的稳态值可能同时具有来自神经元和血管的贡献。
正如在电生理学中所观察到的,持续的刺激也可以引起一些神经元群的相反应(见图2和图3)。例如,Harms和Melcher(2003)的研究证明,听觉皮层中相同的fMRI体素可以根据音调频率(表明相反应的神经元起源)做出短暂或持续的反应。模拟结果表明,这些相位反应极有可能是由神经元引起的(见图4)。Bandettini及其同事(2012)证明,在平均大量的实验run后,大脑的大部分都会对刺激做出反应,许多体素表现出onset和offset反应。因此,相同的物理刺激可能会引起不同类型的反应。在模拟BOLD信号时,必须考虑给定刺激范式下不同的神经元反应。
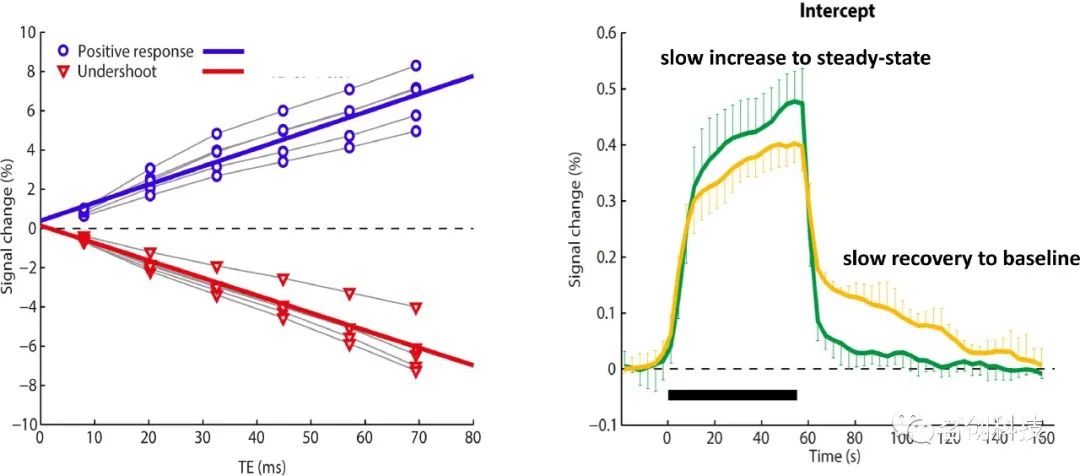
图3
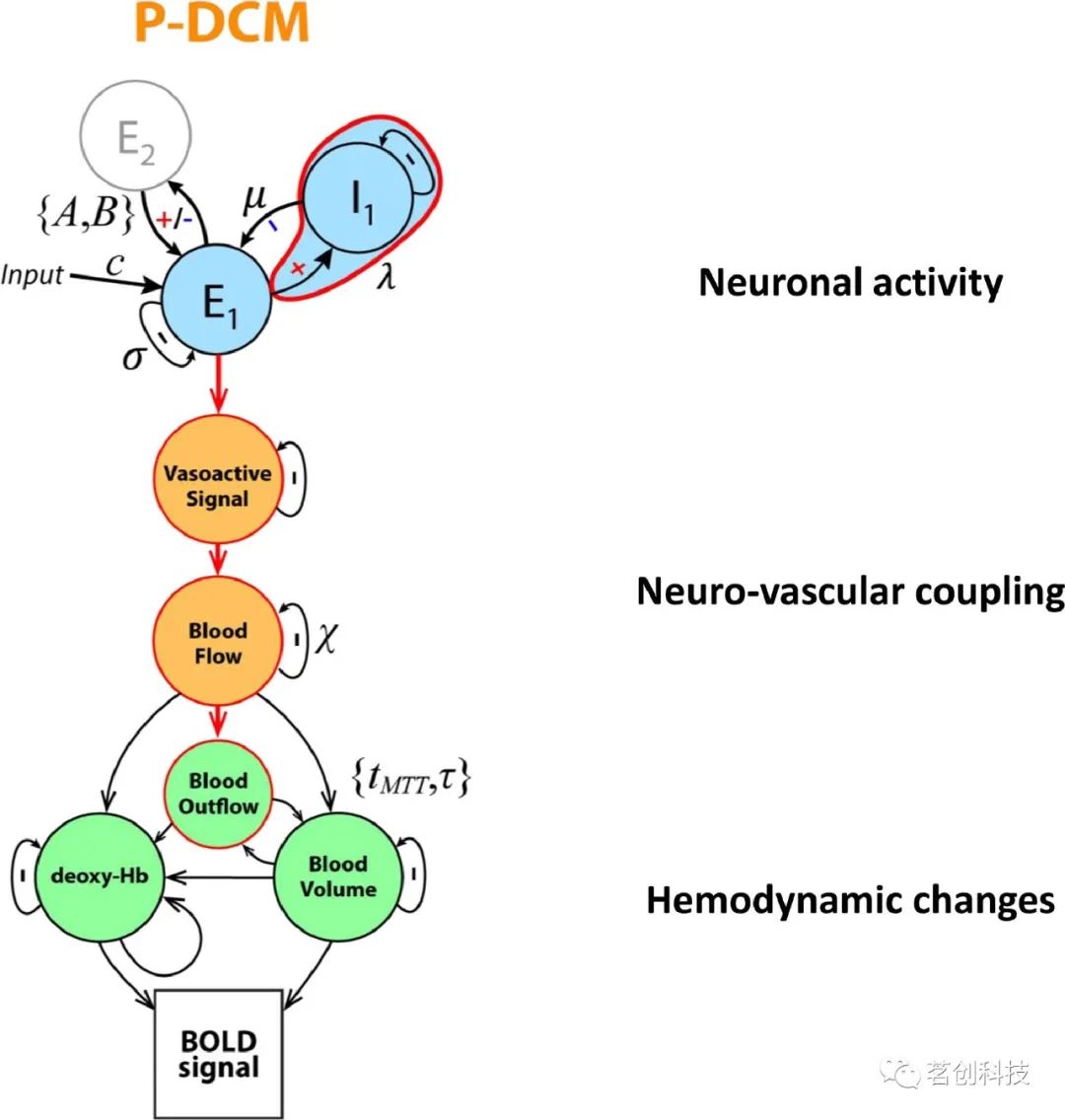
图4.生理信息动态因果模型(P-DCM)的建立。
刺激后下冲
一个常见的现象是(尤其是在感觉区域),BOLD信号在刺激结束后下降到基线以下,称为刺激后下冲(“fMRI中最具争议的问题之一”),这可能需要较长的时间才能恢复到基线(>30s)(见图1)。众所周知,BOLD信号在刺激后下冲的持续时间和幅度取决于刺激类型和持续时间。值得注意的是,在视觉区域V1中(作为对闪烁棋盘刺激的主要反应区域),某些受试者的刺激后下冲几乎与刺激期间的正向BOLD反应一样大(参见图5底图和图3)。
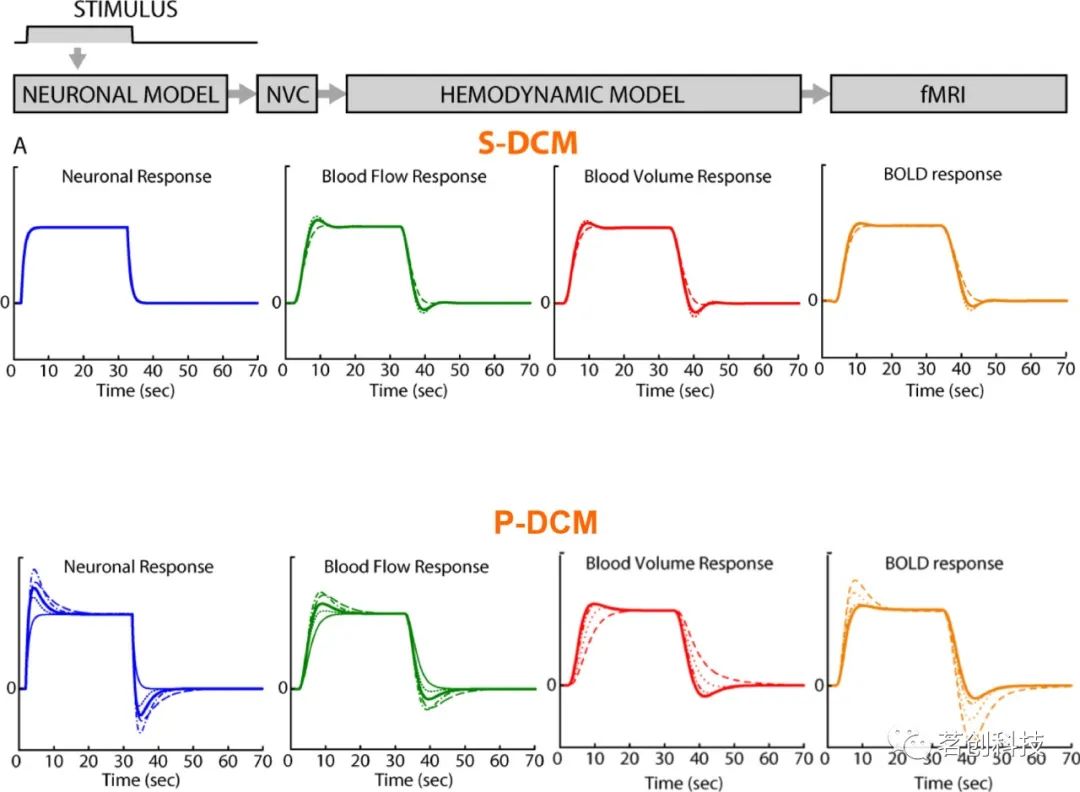
图5.来自S-DCM(顶部)和P-DCM(底部)block设计所产生的CBV、CBF和BOLD信号的时间进程。神经元和血管参数的变化对S-DCM的BOLD信号影响不大,但对P-DCM的BOLD信号有影响,P-DCM表现出更广泛且生理上更合理的变化范围。
目前对fMRI信号的刺激后下冲有三种不同的、非排他性解释(参见图2):
(1)静脉CBV升高导致每体素的脱氧血红蛋白含量增加:含有大部分dOHb的静脉CBV恢复基线的速度比CBF慢得多。Windkessel-Balloon模型描述了静脉CBV的缓慢演变过程,并基于静脉血管的扩张性,将CBV的长时间调节常数视为血管的生物力学特性。
(2)氧合血红蛋白的输送减少:CBF在刺激终止后降至基线以下,这可能是由于刺激后神经元抑制或兴奋-抑制平衡活动的减少所致。CBF假说认为fMRI信号的下冲具有神经元起源,这与动物的侵入性电生理观察相一致:动作电位和局部场电位通常在刺激结束后降低。神经元活动的时间进程以及CBF对fMRI信号下冲的贡献是可变的,因此反映的是活跃的组织过程,而不是继发性血管动态。因此,下冲可以提供与刺激期间主要的正向fMRI反应不同的大脑功能信息。
(3)脱氧血红蛋白的持续产生:CBF先于CMRO2恢复至基线水平;也就是说,在刺激期结束后,氧气仍然被消耗。因此,氧摄取超过刺激前的基线水平,产生短暂的缺氧。这也是一种生理上合理的情况,因为健康受试者的神经血管耦合在很大程度上独立于氧代谢,因此,血流动力学和神经元活动的代谢反应原则上可以解耦联。然而,目前只有间接的MRI数据,而没有直接的氧代谢动态数据来支持这一假设。因此,刺激后CMRO2升高的概念可用以替代CBV持续升高的解释或CBF失活作为fMRI下冲的原因。
总而言之,BOLD信号生成模型必须捕获以下实验观察结果:
①神经元活动(即fMRI背景下的局部场电位(LFP))的时间进程可以根据刺激和脑区的不同而具有不同的形状。对于block设计实验,适应性和刺激后抑制引起的初始过冲通常是LFP的观测特征。
②由于balloon效应,静脉中的CBV在刺激期间增加,而在刺激后下降的速度比CBF慢得多。这与长时间刺激尤其相关。因此,在短时间刺激下,静脉中的CBV变化较小,并且总的CBV变化主要由毛细血管前阻力血管主导。
③BOLD信号的初始过冲和刺激后下冲可以同时具有神经元和血管的贡献。这两种贡献都随刺激特征(例如刺激持续时间和强度)的变化而变化。
最近引入的P-DCM是一种DCM变体,它可以模拟这些观察结果(即可能与神经元和血管有关的可变神经元反应、balloon效应、BOLD信号过冲和下冲)。它基于质量守恒原理,由图4所示组成:(1)该模型将局部神经元群分为兴奋性和抑制性神经元亚群,该模型中的兴奋性时间过程模拟了观察到的LFPs;(2)将兴奋性神经元活动和CBF变化联系起来的神经血管耦合(NVC)作为前馈机制;(3)采用balloon模型来描述血流动力学变化(即CBF-CBV解耦);(4)BOLD信号方程将这些生理变化转化为自旋和梯度回波fMRI信号,提供了不同的场强,最高可达16.4T。
新血流动力学模型对有效连接的影响
P-DCM的每个生理组成部分(即兴奋-抑制神经元模型,前馈NVC和balloon效应)都可以独立地纳入或排除在生成的BOLD信号模型中,因此可以与神经元和血管过程的替代描述相结合(例如,P-DCM中的前馈NVC可以与反馈NVC互换以创建DCM的新变体)。研究者通过3T视觉运动实验,考察了P-DCM的哪些组成部分相对于之前的DCM(即原始DCM和双神经元状态DCM(分别为S-DCM和2S-DCM))有改善。结果发现,P-DCM的所有生理组成部分在所有ROI的BOLD信号时间进程中都有更好的拟合。在另一项研究中,研究者证明了P-DCM框架可以成功地应用于从人类或动物中获得的其他多模态数据(例如,同步采集的CBV和BOLD数据,或同步采集的LFP和BOLD数据)。
错误建模BOLD信号瞬态的影响
Havlicek等人(2017c)研究了P-DCM确定的有效连接模式,并将其与S-DCM(和2S-DCM,此处未显示)获得的模式进行了比较。要求受试者相对于左侧或右侧半视野刺激进行同侧或对侧手指敲击。在这些条件下,共有五个脑区被激活,然后进一步分析这些脑区与不同DCMs的有效连接。结果表明,P-DCM能够更精确地拟合不同区域的时间进程。此外,S-DCM(和2S-DCM)无法解释BOLD信号瞬态,导致在单一条件下脑区之间的有效连接参数错误(图6)。通过将认知假设表示为有效连接图可以发现,此类错误可能会拒绝实际上由fMRI数据支持的认知假设。对于block设计,S-DCM无法模拟任何显著的初始过冲和刺激后下冲。S-DCM导致连接估计改变的原因是,错误的内在连接值必须考虑到由于时间过程的局部拟合不佳而导致的时间过程中无法解释的方差。例如,V1的时间进程有非常大的下冲,无法通过S-DCM中实现的局部血流动力学模型来拟合。通过假设从SMA到V1的有效连接为负,可以解释V1中的一部分下冲。相比之下,P-DCM中的血流动力学模型能够通过局部神经元和血管动力学完全解释这一现象,因此,不需要额外的SMA到V1连接来准确地拟合数据。
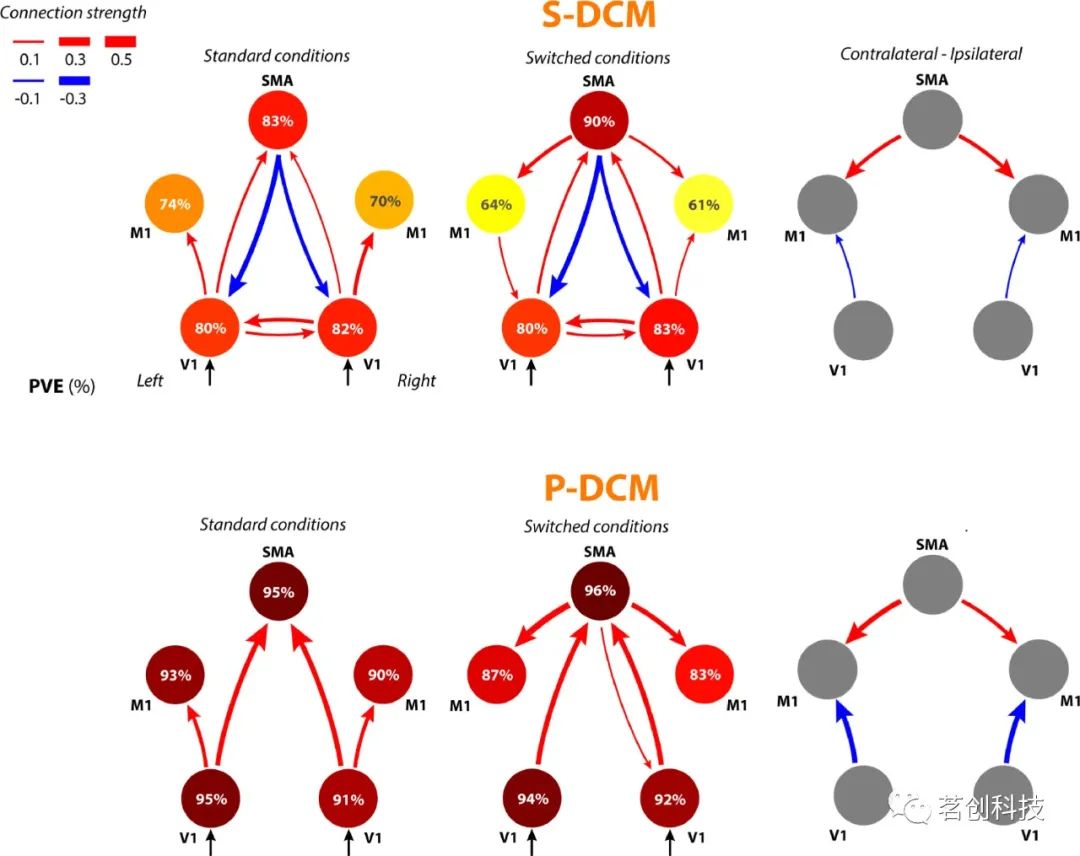
图6.在3T场强下,人类视觉运动实验的有效连接图。圆圈中的数字和颜色表示模型解释的时间过程的方差;红、蓝箭头分别表示正、负有效连接,箭头粗细表示连接强度。
如何知道BOLD信号瞬态是由局部机制诱发,而不是由远距离脑区的反馈诱发的呢?
局部观察到的神经元时间进程既可以由局部过程引起,也可以由其他脑区的远程有效连接引起。因此,没有证据表明P-DCM的兴奋-抑制模型和因果连接图在给定刺激条件下是正确的。重要的是要认识到,统计学框架(如DCM中使用的框架)提供的是支持或反对特定认知或生理假设的统计学证据,而不是证明。这种证明只能通过直接干预或参数记录来提供,例如通过侵入性方法或利用直接刺激(如深部脑刺激或光遗传学)测量轴突的动作电位。然而,S-DCM和2S-DCM不能捕获侵入性电生理学观察到的LFP时间进程,也不能捕获血流动力学观察结果,如CBF-CBV解耦。因此,目前P-DCM在生理学合理性和统计学证据方面更具优势,应该作为DCM中的生成BOLD信号模型以及与DCM共享相同生理模型的建模方法,例如虚拟大脑。
层状BOLD信号模型
梯度回波BOLD信号主要由皮质内上行静脉和软膜静脉的信号组成。因此,BOLD信号也可以在没有神经元活动变化的位置被检测到。在大静脉流经的区域,该区域的神经元活动可能会被掩盖,甚至被错误地归因于该区域。其中两个突出的例子是V4和杏仁核。因此,检查这些区域神经元过程的研究在解释梯度回波fMRI数据或者依赖不易出现此类伪影的MRI序列时必须谨慎。在基于ROI的DCM中,通常会忽略相邻体素的空间依赖性,因为这些体素大部分都包含在同一个ROI中。然而,对于跨皮层深度检测认知过程的高分辨率fMRI,BOLD信号的空间特性不可忽视。
考虑到皮质内上行静脉对皮层表面的单向空间模糊,研究者提出了一个动态层状BOLD信号血流动力学模型(图7)。与P-DCM类似,皮层深处的BOLD信号有来自局部微血管的贡献。此外,dOHb在上行静脉中的变化,将血液从各层输送到软脑膜静脉,就会引发BOLD信号,而BOLD信号又会在神经元活动区域上方的各层传递。因此,在高分辨率梯度回波fMRI中,皮层深处的BOLD信号有来自局部微血管和上行静脉的贡献。该模型基于质量守恒原理,用于确定跨皮层深度的空间和时间点扩散函数。点扩散函数允许对测量到的BOLD信号进行“去卷积”,以获得不受上行静脉影响的层间BOLD信号分布。
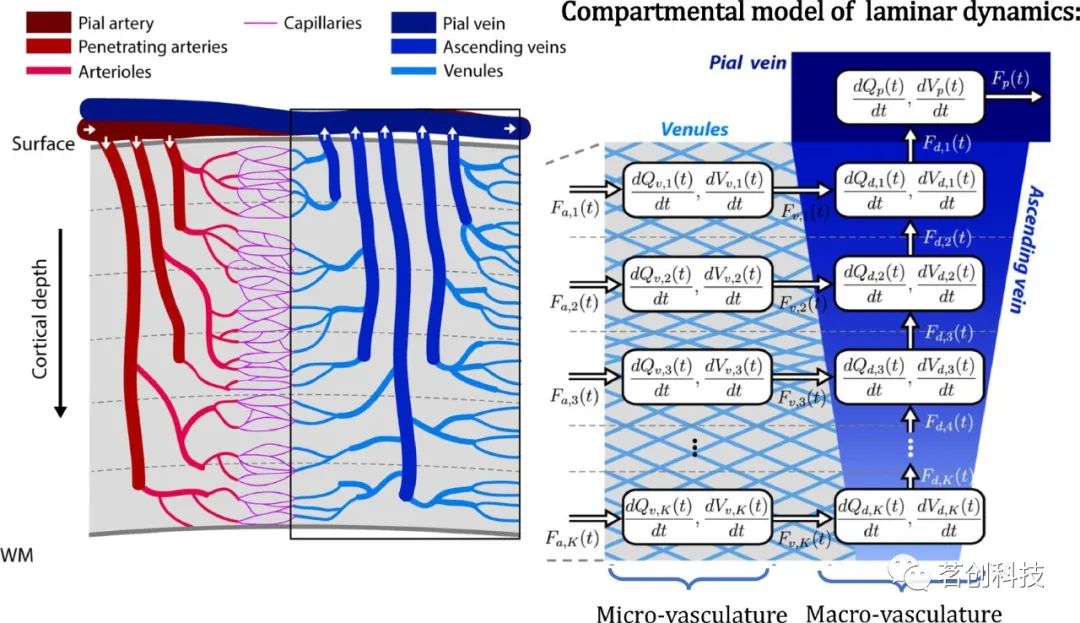
图7.动态层状BOLD信号血流动力学模型。
P-DCM的局限性和未来发展方向
标准分辨率P-DCM和层状BOLD信号模型是确定标准分辨率和高分辨率BOLD信号数据中局部神经元活动和远程有效连接的强大工具。它们可以提供关于空间和时间BOLD信号观测的神经元和/或血管方面的解释。此外,P-DCM与先前的DCM变体一样存在一些局限性,主要包括(但不限于)以下几点:a)对于复杂的自然刺激(如观看电影),有效连接可能会在run中发生变化;b)随着ROIs数量的增加,可能的连接数量会呈指数级增长,计算时间也会大大增加,因此DCM研究中所考察的ROIs数量受到了限制(通常<6个ROIs);c)通常情况下,ROIs在代表特定认知假设的图形模型中彼此相关;然而,在一个ROI中,一部分脑区的体素子集或神经元亚群可能与其他脑区具有不同的有效连接性。因此,需要确定并考虑ROI内独立计算单元数量的探索性方法;d)P-DCM尚未应用于静息态数据;原则上,用于确定静息态有效连接的随机或频谱DCM的统计框架可以与新的P-DCM方法轻松结合;e)P-DCM中的神经元模型尚未得到同步电生理和fMRI数据的充分验证。
此外,层状BOLD信号模型的另一个局限性是,每个皮层深度都使用独立的兴奋-抑制模型进行建模。然而,兴奋性和抑制性神经元覆盖并分支在几层中。因此,即使MUAs和LFPs主要分别由局部动作电位和膜电位产生,但建模必须考虑神经元的柱状和层状环路。所谓的典型微环路模型被认为代表了新皮层中局部神经元群的回路,并且可以很容易地与P-DCM的其他组成部分(即局部前馈NVC和balloon效应)相结合。描述层状神经元模型也是探索新型层状BOLD信号建模的一种方法。然而,为了测试这种方法相对于P-DCM的兴奋-抑制神经元模型的有效性和统计优势,需要对电生理学和BOLD信号进行高时空测量。可以想象的是,典型微环路模型不一定适用于整个大脑。未来可能会使用基于组织学数据(如BigBrain),以及神经递质及其分布的启发式方法推导出特定于脑区的微环路模型。最后,生成BOLD信号模型的生理参数可能会因患者而异,因此,在与健康受试者进行组间比较时,这些差异可能会对有效连接参数甚至是一般线性模型(GLM)获得的统计图产生影响。
综上所述,BOLD信号是对神经元活动的间接反映,其受到外源输入、局部加工和远程有效连接的影响。由于神经元活动通常是认知神经科学感兴趣的生理过程,如果直接在BOLD信号水平上分析数据,而不是在分布式神经元活动水平上进行分析,神经血管耦合、血流动力学和BOLD信号物理学等生物学特性可能会导致错误的结论。鉴于BOLD信号可能并不真实地反映神经元活动模式,令人惊讶和遗憾的是,使用fMRI的计算神经科学领域并未利用(极少数例外)BOLD信号的生物物理模型进行模型反演。
然而,模型反演有望从伴随的BOLD信号中确定神经源。BOLD信号的生理模型原则上必须是可逆的,即在准确反映主要生理过程的同时,自由参数的数量必须尽可能少。这就要求BOLD模型在宏观层面上描述生理过程,而VANs则相反,其在fMRI数据的自由参数数量非常多且空间分辨率相对较低,因此它们是不可逆的。此外,BOLD信号前向模型的分量必须符合侵入性和非侵入性实验数据。P-DCM与S-DCM的比较结果表明,在自由参数数量几乎相同的不同生理假设条件下,P-DCM对BOLD数据的拟合效果更好。因此,P-DCM在统计学上更具优势,因为它能够覆盖更大范围的实验观测BOLD信号时程。此外,为了使用模型反演来区分神经元和血管参数,必须仔细选择实验设计。最后,值得注意的是,DCM框架不是一个用于数据探索的工具,而是用于认知假设检验的工具,即统计结果对某些假设提供更高的证据,但并不声称生成或提供“正确”的假设。合理假设的生成是一门艺术性工作,需要整合先前的理论概念和实验数据。因此,必须谨慎解释DCM(以及整个fMRI)的结果,并且解释的指导原则应该是汇集许多不同神经科学实验的证据,最好是来自不同成像模态的证据。
原文:Kâmil Uludağ, Physiological Modeling of the BOLD signal and Implications for Effective Connectivity: a primer, NeuroImage (2023), doi: https://doi.org/10.1016/j.neuroimage.2023.120249