内容一览:基因渐渗与葡萄的驯化、遗传改良密切相关。先前研究揭示了欧洲栽培葡萄中,野生葡萄基因渐渗的基因组信号,但尚未深入研究这些渐渗事件发生的时间、方式、基因组模式和生物学效应。本文中,中国农业科学院深圳农业基因组研究所的研究人员,使用机器学习的种群遗传分析方法,对栽培葡萄以及野生葡萄进行了重测序数据分析,这对于葡萄育种具有重要意义。
关键词:基因渐渗 葡萄驯化 机器学习
本文首发自 HyperAI 超神经微信公众平台~
基因渐渗是指基因从一种物种或种群,渗入到另一种物种或种群的过程。它通常发生在不同物种之间的杂交和回交过程中。在基因渐渗中,外源基因通过杂交进入目标物种的基因池,与目标物种的本地基因发生混合。这种渐渗可以导致目标物种的基因组发生改变,获得新的遗传变异和多样性。
葡萄的基因渐渗是指野生葡萄 (Vitis vinifera ssp. sylvestris) 与栽培葡萄 (V. vinifera ssp. vinifera) 之间基因交流和基因流动的过程。在自然界和人工选择的影响下,野生葡萄与栽培葡萄之间发生基因交换,导致栽培葡萄中出现野生葡萄的遗传特征。
近期,中国农业科学院深圳农业基因组的研究人员,研究了驯化葡萄与其欧洲野生近缘种之间的渐渗历史,他们使用基于机器学习的群体遗传方法,揭示了葡萄风味的形成机制,对其基因特征及其对葡萄育种的影响提供了深入的见解。目前,该研究成果已发表在《Proceedings of the National Academy of Sciences》期刊上,标题为《Adaptive and maladaptive introgression in grapevine domestication》。

该研究成果已发表在《Proceedings of the National Academy of Sciences》期刊上
论文地址:
https://www.pnas.org/doi/abs/10.1073/pnas.2222041120
数据集
研究人员对一组 345 个测序样本进行了分析,这些样本包括酿酒葡萄和食用葡萄,涵盖了野生葡萄的广泛地理分布范围,其中包括来自欧洲的 72 种野生葡萄 (V. vinifera ssp. sylvestris)、来自中东和高加索地区的 36 种野生葡萄和 231 种驯化葡萄 (V. vinifera ssp. vinifera),以及代表北美外群物种的 Vitis californica (n = 3) 和 Muscadinia rotundifolia (n = 3)。此外,研究人员还对欧洲野生葡萄进行了采样以覆盖欧洲和近东的预测分布区域。
数据 (基因组序列;script) 已存放在 GitHub 中:
GitHub - zhouyflab/Grapevine_Adaptive_Maladaptive_Introgression: Adaptive and maladaptive introgression in grapevine domestication
实验结果
基因渐渗历史
首先,研究人员评估了欧洲野生葡萄、酿酒葡萄和食用葡萄之间的差异,以深入了解种群和驯化历史。值得注意的是,欧洲野生葡萄 (EU sylvestris) 形成了一个独特的群体,与酿酒葡萄共享少数混合成分。
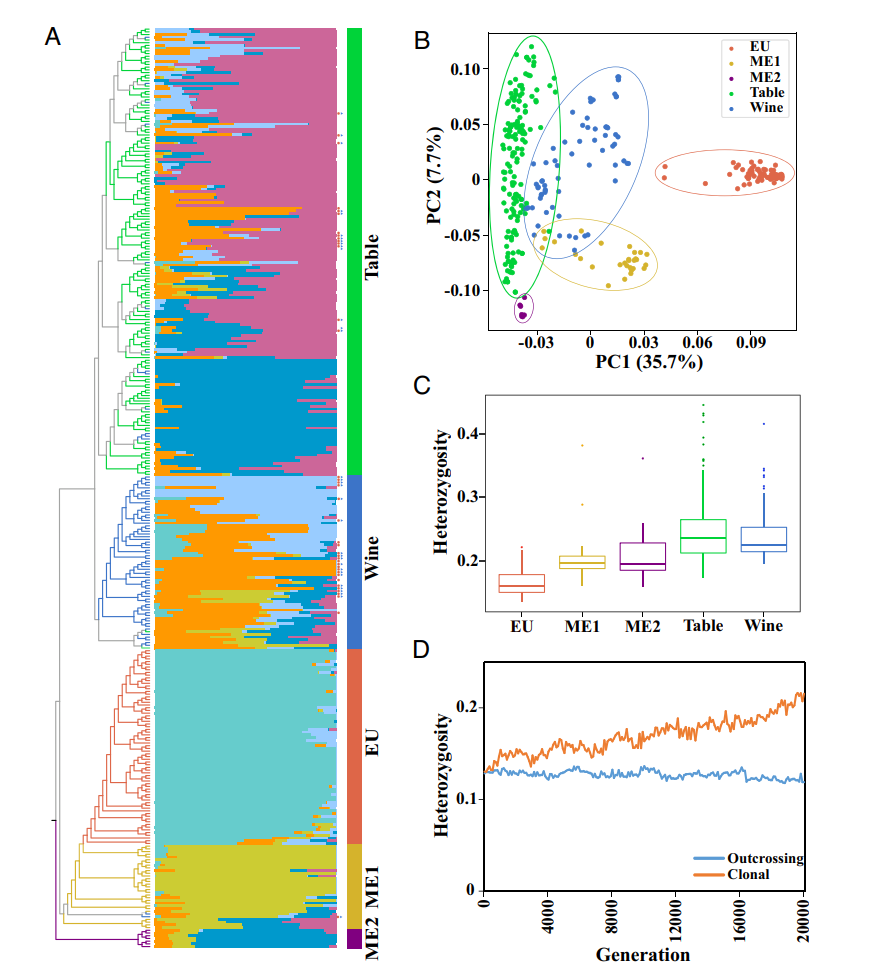
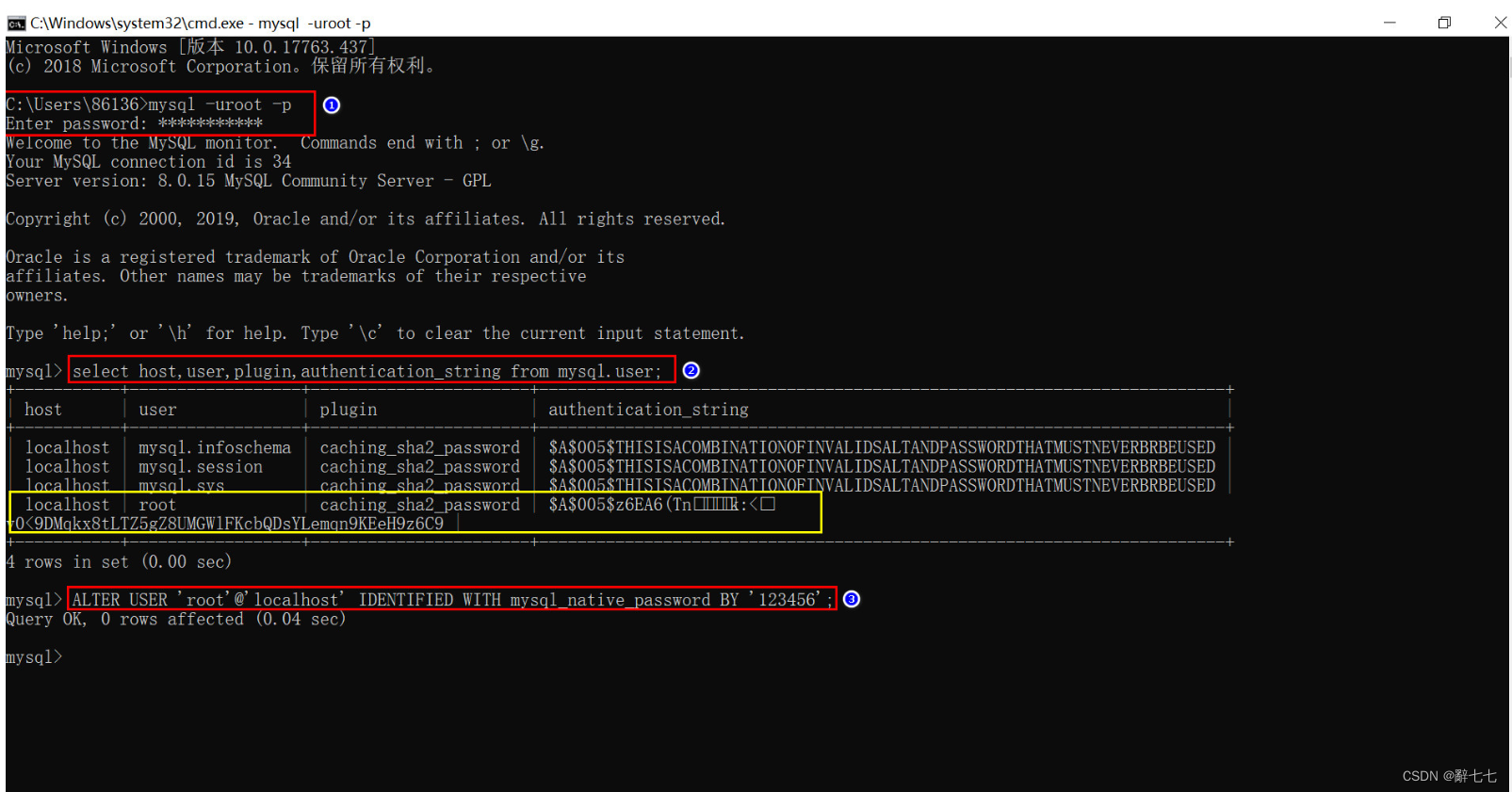
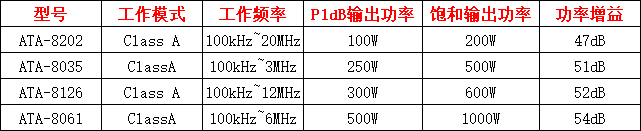
图 1. 不同葡萄品种对比
A:混合分析的系统发生树。
在系统发生中,分支的颜色反映了不同的群体:ME 1,黄色;ME 2,紫色;野生葡萄,红棕色;酿酒葡萄,蓝色;食用葡萄, 绿色。Admixture 图, K = 6 。图右侧的红点和蓝色三角形分别显示来自食用或酿酒组的叶绿体或线粒体是否明显起源于欧洲野生葡萄。
B:五个组的 PCA。
C:五组的杂合性。
D:不同传播类型下的正向模拟结果。蓝线表示异交繁殖,而橙色线表示克隆繁殖。
图 1 显示,酿酒葡萄和食用葡萄在进化过程中发生了早期的分化,表明它们在用途上有明显的区别。野生葡萄样本也呈现出单系群,但分为三个不同的群体:欧洲野生葡萄(EU)、位于里海周围葡萄 (ME 1) 和位于地中海附近的肥沃新月地带的葡萄 (ME 2)。通过 主成分分析 (PCA) 和祖先成分比例的估计,也证实了这些种群之间的分化。酿酒葡萄和食用葡萄 (均为 0.24) 的杂合性比野生种群更高 (欧洲为 0.17,ME1 为 0.20,ME2 为 0.22),这可能是由于历史上的渐渗事件和长期的无性繁殖导致了杂合突变的积累。
基因渐渗方向
研究人员在最初的模型基础上,估计了食用葡萄、酿酒葡萄和欧洲野生葡萄之间可能存在的 34 种基因流动模式。根据最佳模型,fastsimcoal 推断欧洲野生葡萄大约在 4 万年前发生了分化。
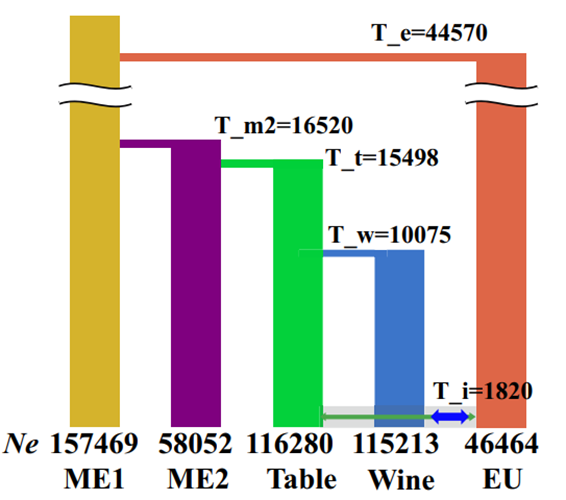
图 2:渐渗检测
水平分支中的时间 (T) 是每个组的分化时间 (年),T_i 表示推断的渐渗开始时间。
底部的数字是每组的估计有效种群规模 (Ne)。
图 2 显示,在最佳模型下,fastsimcoal 推断出 EU 类群在大约 4 × 104 年前分化;食用葡萄的驯化早在 1.5 × 104 年前就开始了;大约在 1.0 × 104 年前,酿酒葡萄从食用葡萄中分离出来。模型表明,欧洲野生种群和驯化种群之间的基因流动始于 1.8 × 103 年前。此外,最佳拟合模型还表明,基因从欧洲野生种群流动到驯化种群的概率很高,向酿酒葡萄 (1.7 × 10−4) 的迁移率是食用葡萄 (3.8 × 10−5) 的 5 倍。
基因渐渗区域
研究人员使用机器学习方法来识别葡萄基因组的基因渐渗区域,以及这些区域的基因组特征。
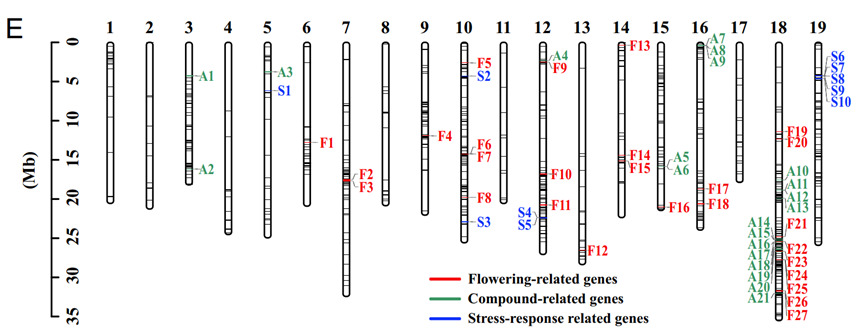
图 3:渐渗区域的三种基因
Filet 预测的假定基因渐渗区域用黑线标记在 19 条染色体上。
三种颜色代表三种基因渐渗区域内的基因
红色:开花相关基因
绿色:芳香化合物相关基因
蓝色:应激反应基因。
研究人员对整个基因组中的渐渗选择基因进行了 GO 功能研究,并重点关注开花相关基因、风味相关基因和应激反应相关基因。图 3 显示,许多与开花相关的基因在渗入后选择,其次是芳香族化合物相关的基因。大多数 GO 分类的富集也与风味相关,包括木质素降解过程、L-苯丙氨酸降解过程和肉桂酸生物合成过程。因此,研究人员推断酿酒葡萄和欧洲野生葡萄之间的适应性渐渗主要影响了与风味相关的性状。
为了解等位基因的渐渗动态,研究人员对异交繁殖和克隆繁殖的渐渗进行了正向模拟。

图 4:非渐渗区域和渐渗区域中有益 SNPs 和有害 SNPs 的 SFS
E:整个渐渗群体中渐渗等位基因的总数
F:整个渐渗群体中不同类型的渐渗等位基因的数量
G:异交组第 500 代渐渗有益和有害等位基因的 SFS
H:克隆组第 500 代渐渗有益和有害等位基因的 SFS
研究发现,杂交后渐渗等位基因 (包括有益和有害等位基因) 的总数都有所增加,直至达到平衡。这些模拟表明,在克隆和异交系统中,渐渗等位基因的情况可能存在显著差异。渐渗对有益和有害变异的补充起到了重要作用,这些变异可能成为葡萄栽培基因组设计的主要目标,包括在有性繁殖过程中清除潜在的有害变异。
AGIS:专注农业科技创新
值得注意的是,该论文的多名作者均来自中国农业科学院深圳农业基因组研究所。基因组所成立于 2014 年,通过整合生物学和大数据科学,来认识与利用农业生物基因组,服务全球农业生产。基因组所的长期愿景是致力于通过颠覆性创新来促进全球农业可持续发展,服务于个性化食品供给体系,并提升人类健康水平和农民社会地位。
基因组所在包括 Science、Nature、Cell 等顶级期刊在内的杂志上发表 SCI 论文 620 多篇,农业基因组学等研究领域占据世界前沿。除了研究葡萄的基因渐渗问题,该所还发布了另外两篇与葡萄基因相关的研究成果,不仅联合国内外学者首次绘制了葡萄完整基因组图谱,还揭示了野生葡萄的抗皮尔斯病的全基因组效应和气候适应性机制。
所长表示,基因组所还是一个非常年轻的科研院所,建设国际一流的农业科研院所任重而道远,将立足新时代新征程新使命,以坚持「四个面向」助力高水平科技自立自强,为建设「农业强国」继续做出我们的贡献。目前,基因组所联合深圳市相关部门提出了建设「深圳国际食品谷」建议,规划已得到市政府批复,将在深圳构建农业食品产学研协作生态,做出科技推动农业食品产业转型升级的先行示范。
基因组所官网:
中国农业科学院农业基因组研究所
参考文献:
[1]https://www.163.com/dy/article/I6KVQLV205328VPM.html
[2]https://www.sohu.com/a/682674856_121124027
[3]https://www.caas.cn/xwzx/kyhd/60f2e9b4dff84bed9e315b7097aeb26b.htm
[4]https://www.agis.org.cn/bsgk/yjsjj/index.htm
[5]https://www.agis.org.cn/xwzx/kyjz/677aecae97c448c9bed7e89f95daae7f.htm