改变微生物群落可以增强树木对气候变化的耐受性
Shifting microbial communities can enhance tree tolerance to changing climates

Research Article,2023-5-25,Science, [IF 63.714]
DOI:10.1126/science.adf202
第一作者:CASSANDRA M. ALLSUP
通讯作者:RICHARD A. LANKAU
作者单位:University of Wisconsin–Madison
- 摘要 -
气候变化使得物种拥有了超出其进化的耐受性。植物种群必须适应、调整或改变生长地方以避免灭绝。然而,由于植物与塑造其表型的不同微生物群落相关联,微生物群落的变化可能会影响植物对气候的耐受性。我们研究表明,当面临干旱、高温或低温压力时,接种了来自更干燥、更温暖或更寒冷地区的微生物群落的树苗分别显示出更高的存活率。微生物介导的耐旱性与假丝酵母真菌的多样性增加有关,而耐寒性与较低的真菌丰富度有关,可能反映了非适应性真菌的减少。了解微生物介导的气候耐受性可能会有利于我们预测和管理森林生态系统对气候变化的适应性。
- 前言 -
气候变化超出了种群的气候耐受范围,据估计,在下个世纪面临灭绝的物种高达16%。当面临不断变化的气候时,物种必须迁移到最佳气候,或适应新的气候条件;否则,它们将面临灭绝。然而,物种存在着多样化的群落,物种间的相互作用可能提供了适应气候的其他途径。特别是,宏观生物与不同的微生物群落(即它们的微生物组)存在着很多联系,这些群落影响了它们的表型和适应气候变化等的性质。
植物与微生物的关系不断改变以满足基本需求。根茎层和内生微生物经常提高植物对冷、热和干旱的耐受性。特定的微生物群对植物种群适应极端环境至关重要,来自特定环境的微生物群落也可以影响人工条件下植物对环境压力的反应。几乎所有树种都与丛枝菌根(AM) 或外生菌根 (EM) 真菌关联,它们代表了系统发育和功能上不同的共生体。菌根真菌可以提高植物对许多环境条件的耐受性,因此微生物很大可能介导了植物对气候耐受性。这种有“适应压力”的微生物群落可以通过增加能够在压力下表现更好的微生物菌株或排除不能耐受压力的有害微生物类群来影响植物性能。如果和微生物的联系有助于植物气候生态位,那么我们理解当前和预测未来植物分布的能力可能是不完整的。然而,在自然条件下,不同气候的微生物群落变化在多大程度上可以导致宿主植物的耐受性增强还没有得到验证。微生物群落可能比它们的宿主植物尤其是树木适应得更快以及可能分散的更远。因此植物群落可能通过微生物的适应或改变来抵御气候变化。
气温升高和干旱会通过物种丰度的变化、物种的重新分布以及森林向其他生物群落的转变重塑森林,并可能对全球气候系统产生影响。为了研究微生物群落的地理变化如何影响树木对气候压力的反应,我们首先收集了来自不同温度和降水梯度的 12 个位置的微生物群落。然后,我们研究了引入这些群落如何影响树苗在扩大和缩小范围的边缘位置以及对气候压力的反应。在每个地点,我们在环境和降雨减少的条件下种植了几个树属的幼苗,包括那些与 AM 或 EM 真菌相关的树种。在这两个地点和降雨处理中,这导致幼苗经历了冷、热和干旱的变化。通过接种微生物群落,可以提高幼苗的存活率,这些微生物群落来自与每个田间地点或实验处理中幼苗面临的主要气候压力最匹配的石灰质地区。微生物的耐热性和耐旱性仅发生在与 AM 真菌相关的树木中,而耐寒性则发生在所有的菌根类型中。这些结果表明,土壤和根部微生物群落可能是森林的气候耐受性的另一个来源。
- 结果 -
匹配微生物群落和气候压力可以提高野外条件下的树苗生存率
在北部地点(图1),我们种植了混合的树种,这些树种要么是该地点的原生树种(但其分布中心在该地点以南),要么是该地点以南的原生树种(表S1)。这个实验模拟了范围扩张的过程,在这个过程中,种群在因气候变化而变得宽松的地区扩张或建立。我们预计,越冬生存将是该地幼苗的主要气候压力。因此,我们预测,当预先接种来自寒冷地区的微生物群落时(微生物诱导的耐寒性),幼苗的存活率将达到最大化。支持这一假设的是,在环境降雨条件下,接种了来自冬季较冷地区的土壤微生物的幼苗具有较高的3年存活率[系统发育广义线性混合模型 (PGLMM),P = 0.031;图 2A和表S1 ]。当幼苗在降雨量减少的条件下生长时,这种影响不明显(PGLMM,P = 0.823;图2B和表S2)。微生物来源地的最低温度比平均或最高温度更能预测幼苗的存活。微生物来源的干旱度与幼苗的存活率没有明显的关系(表S2)。存活率独立地受到初始幼苗高度的影响(表S2)。这些结果在与AM或EM真菌相关的幼苗之间没有差异(图2A和B,以及表S3)。
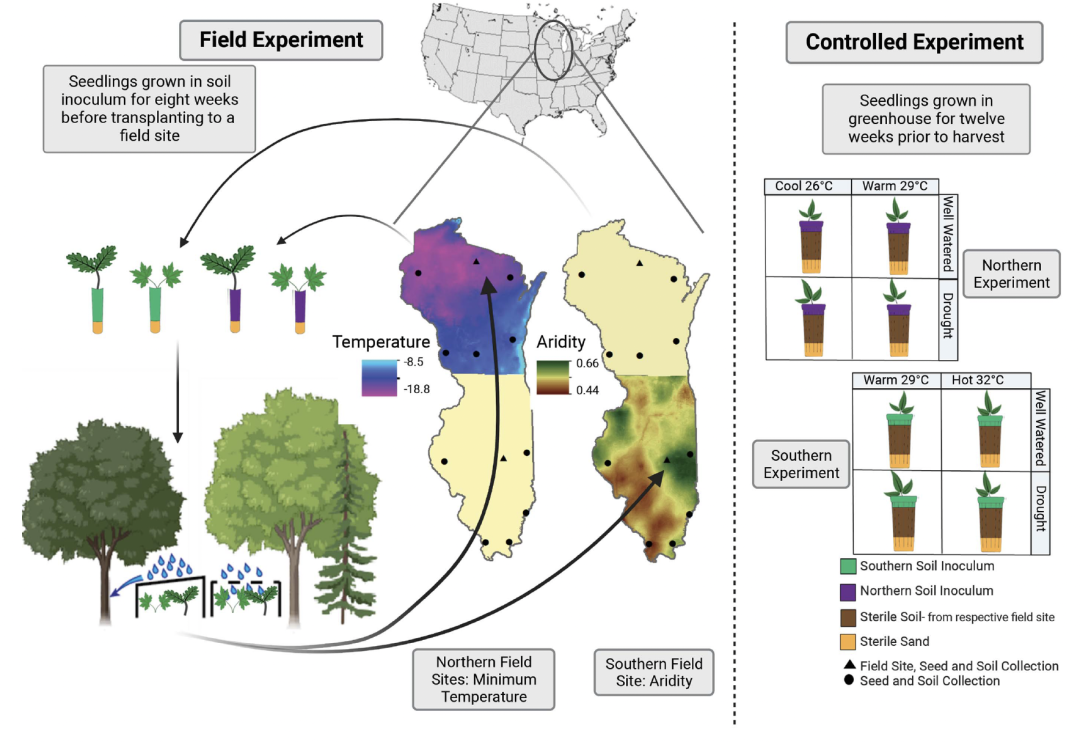
图1. 田间和温室的实验设计。
对北部地区和土壤进行平行实验用来模拟扩张,对南部地区和土壤进行平行实验用来模拟范围收缩。(i)从每个州的六个地点收集活土。(ii)幼苗在无菌条件下发芽,然后在移植到实验地点前用田间活土生长8周(三角形符号)。(iii)两个队列(2018 年和 2019 年种植)的幼苗在降雨量减少或模拟(环境条件)的棚室中生长3年,并按季节监测存活率。(iv) 来自同一地点的活土接种花盆用于控制环境变量,花盆里装的是来自田间实验地点的灭菌土壤。(v) 幼苗在两种温度条件和两种浇水频率下生长12周。温室实验重复3次,每年一次,持续3年,每次重复采集新的土壤。
为了研究微生物如何影响整个3年实验的存活率,我们比较了不同季节的存活率["生长季":春季到秋季的存活率; "越冬期":秋季到春季的存活率]。在第一个夏天,在环境和降雨量减少的条件下,接种来自温暖地区的微生物群落的幼苗存活率最高(PGLMM,分别是P = 0.064和0.031;图2,C和D,以及表S4)。然而,在第一个冬季期间,这种模式发生了逆转;接种了来自较冷地点的微生物群落的幼苗的越冬存活率明显较高(PGLMM,环境条件下的P <0.0001,降雨量减少条件下P = 0.019;图2,C和D,以及表S4)。在第二个夏天,自然环境条件下的存活率与降雨量减少条件下的存活率出现分歧:自然环境条件下的存活率加强了前一个冬天的趋势,而降雨量减少条件下的存活率则抵消了这一趋势(PGLMM,降雨处理×微生物最低温度的相互作用,P=0.025;图2,C和D,以及表S4)。也就是说,在环境降雨条件下,幼苗面临一个主要的气候压力--寒冷的冬季温度,并且通过预先接种来自较冷地区的微生物群落来优化生存。但在降雨量减少的处理中,幼苗面临多种气候胁迫(冬季寒冷以及夏季干旱),因此没有特定的微生物群落能够优化3年内的累积存活率。
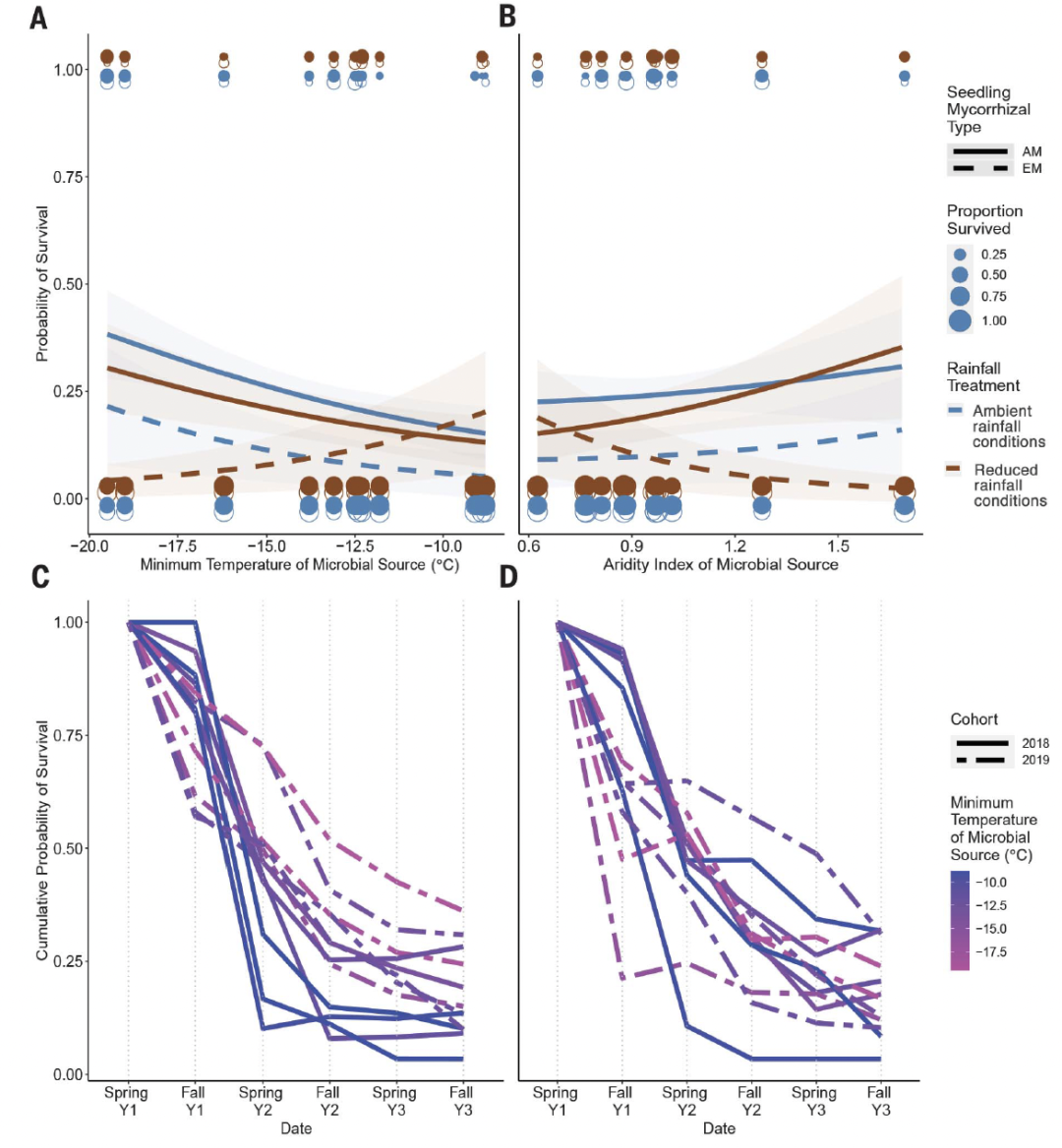
图2. 北部田间的幼苗存活模式与微生物接种源的气候条件的关系。
(AB)存活超过3年的概率与微生物源地的最低温度(A)或干旱指数(B)的关系。干旱指数、降水量/潜在蒸发量:较低的值表示较干燥的条件。蓝色,环境降雨条件;棕色,降水量减少条件。虚线和开放符号,外生菌根(EM)物种;实线和填充符号,从枝菌根(AM)物种。(CD)三年来的累积存活率,按春季和秋季采样日期划分。(C)自然环境降雨条件,(D)降水量减少条件。每条线代表特定微生物接种源的幼苗累积存活概率,根据微生物源地的最低温度着色。实线,2018年的队列;虚线,2019年的队列。
我们选择了伊利诺伊州中部的南部地点,以研究温带树种的高温和干旱压力。这个地点在我们所有目标物种范围的分布中心的南部,对许多物种来说接近南部的分布极限。因此,这个地点代表了一个物种的后缘动态,那里的气候变得过于温暖或干燥,不利于种群的生存(表S1)。在这里,我们预计,在环境条件下,幼苗将主要受到夏季高温的压力。我们假设,当幼苗被接种了来自温暖地区的微生物群落时,幼苗的存活率将得到优化(即微生物诱导的耐热性)。另外,在降雨量减少的处理中,我们预计幼苗将受到更大的干旱压力,因此假设当接种来自更干旱地区的微生物群落时(微生物诱导的抗旱性),幼苗的存活率将得到优化。结果支持我们的假设,在我们的环境降雨处理中,幼苗存活率与微生物来源地的最高温度呈正相关(PGLMM,P = 0.093;图3A和表S5)。在降雨量减少的处理中,通过预先接种来自更干旱地区的微生物群落,存活率得到了极大的提高(PGLMM,P = 0.002;图3B和表S5)。在两个实验地点用Cox回归分析存活时间时,发现了类似的结果(表S2和S5)。
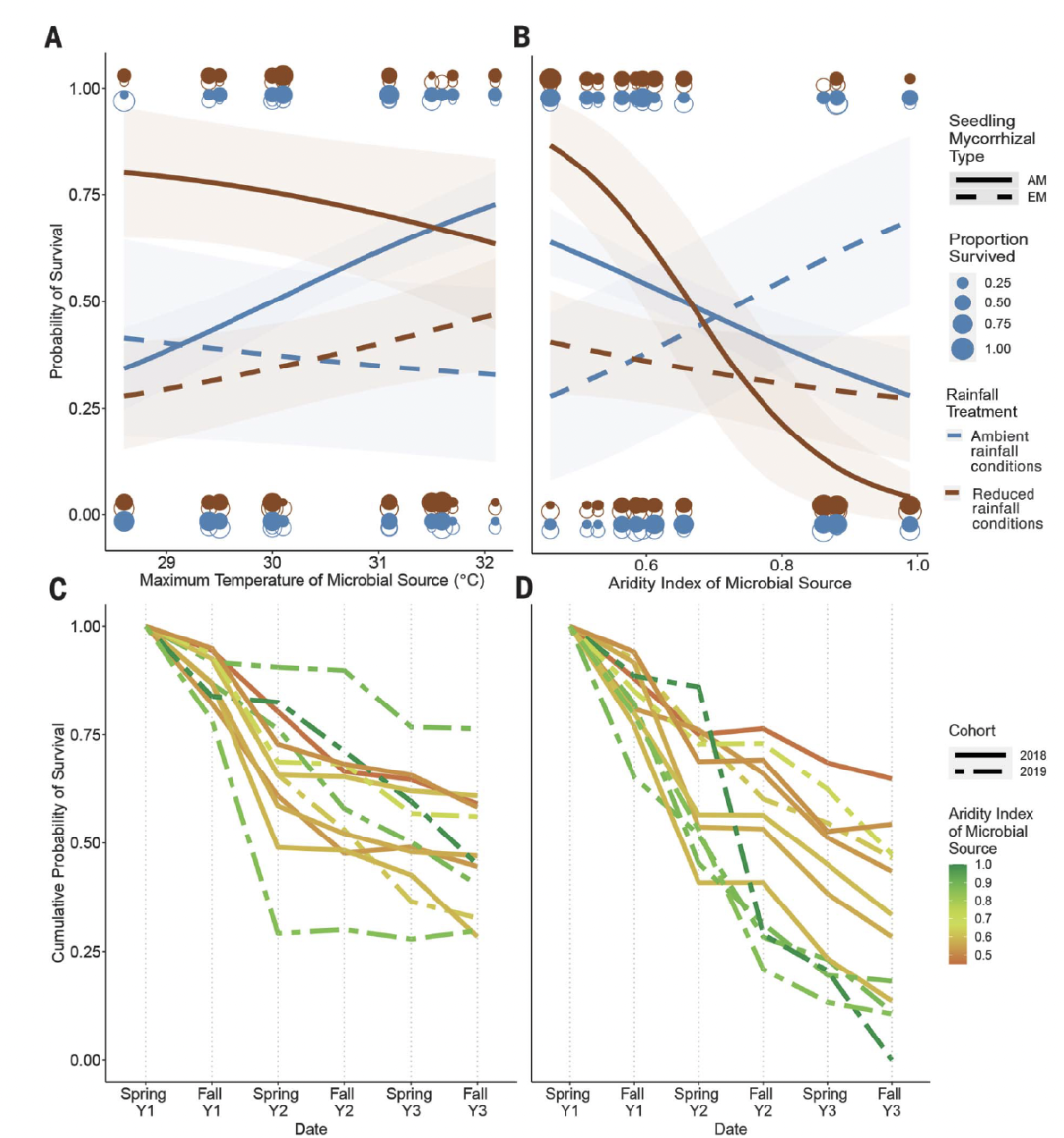
图3. 南部田间的幼苗存活模式与微生物接种源的气候条件的关系。
南部田间地点的幼苗存活率(A和B)。存活超过3年的概率与微生物源地的(A)最高温度或(B)干旱指数的关系。干旱指数,降水量/潜在蒸发量,在土壤采集前的生长季节(5月至10月)测量;较低的值表示较干燥的条件。蓝色,环境降雨条件;棕色,降雨量减少条件。虚线和开放符号,EM物种;实线和填充符号,AM物种。(C和D)三年的累积存活率,按春季和秋季采样日期划分。(C)自然环境降雨条件,(D)降低降雨条件。每条线代表特定微生物接种源的累积存活概率,根据微生物源地的干旱指数进行着色。实线,2018年的队列;虚线,2019年的队列。
与北部地区不同,在我们的南部地区,与AM和EM共生真菌相关的幼苗对微生物接种源有不同的反应。对于与AM相关的幼苗,微生物诱导的气候耐受性的预测得到了强有力的支持:当接种来自更高温度的环境的微生物群落时,AM幼苗的存活率较高,但在降雨条件下未改变(PGLMM,分别是P = 0.009和0.405;图3和表S6),但在微生物来源的干旱方面显示出相反的模式(PGLMM,环境条件下P = 0.059,降雨量减少条件下P < 0.0001;图3和表S6)。在任何条件下,EM相关的幼苗的存活率都没有显示出与微生物来源地气候之间的明显关系(图3和表S6)。微生物来源地最高温度的影响广泛分布于各个季节,在大多数季节呈正相关趋势,但与任何一个季节的存活率均未产生统计学上显着的关联(PGLMM,所有P>0.05)。相比之下,微生物来源的干旱影响集中在第二个夏天。第二个夏天的存活率在很大程度上取决于降雨处理与微生物源干旱之间的相互作用(PGLMM,P = 0.014;表S7),因为在降雨量减少处理组中存在非常显著的关联(PGLMM,P = 0.003;表S7)。因此,预先接种来自更干旱地区的微生物群落,可以增强幼苗在最强烈的干旱期间(夏季降雨量减少)的耐旱性。
微生物介导的控制条件下的耐旱性
我们的田间实验无法将幼苗经历的气候条件与我们地点的其他条件(如土壤性质)分开。此外,我们接种的微生物群落的变化可能是由于来源地之间的许多差异引起的。特别是,在我们北部的一组实验中,与温暖的来源地相比,较冷的微生物来源地的土壤条件也与田间地头的土壤条件更相似(例如,较高的土壤有机质水平)。由于微生物群落可以促进植物生长适应当地的土壤条件,如果我们接种的微生物群落对我们田间土壤的适应性不同,这可能被误认为是微生物诱导的气候耐受性。此外,微生物群落对幼苗健康的直接影响也可能不同。苗木健康也可能受病原体的直接影响而与气候耐受性无关。因此,我们使用每个田间地点的消毒土壤进行了盆栽实验,接种了我们田间实验中使用的相同的微生物群落,在自然环境温度和夏季高温下以特定的浇水频率生长。我们用这个实验来测试在我们的田间观察到的模式是否取决于特定的气候条件,或者更确切地说研究微生物群落、非生物土壤条件和幼苗之间的非气候关系。
在控制条件下,与我们的田间结果不同,我们北部实验点的土壤中的幼苗生长不受微生物接种源气候的影响。这个实验不能直接确认微生物介导的耐寒性模式,因为盆栽实验中的幼苗没有经历过冬季温度。南部地区土壤中的幼苗生长表现出与田间实验中类似的模式。特别是,在接种了来自干旱地区的微生物群落的盆中,幼苗的生长得到了加强,但只有在干旱压力最大的处理中(高温和限制浇水)(PGLMM,P < 0.001;图4A,表S9,和图S2)。微生物介导的抗旱性模式与田间结果一致,与EM相关的幼苗相比,AM的耐旱性更强(图4A和表S10)。
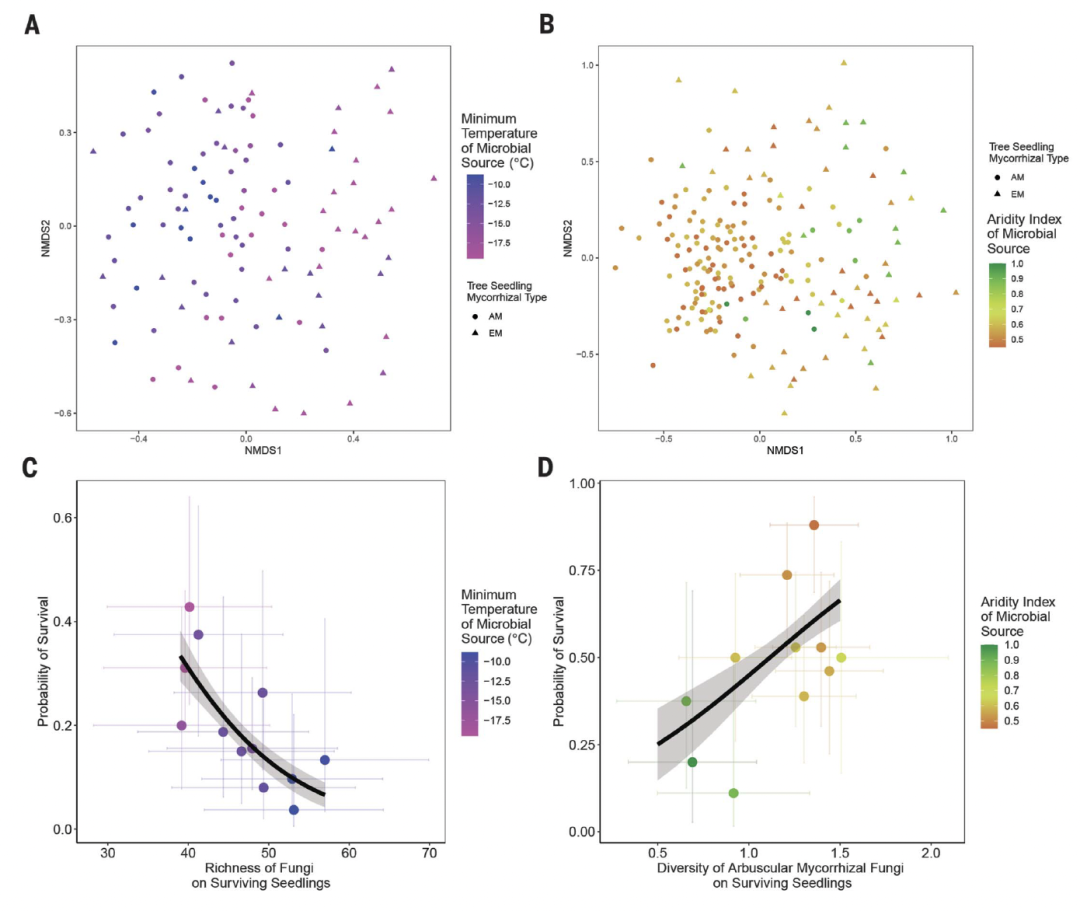
图4. 存活的幼苗上的真菌群落。
最初的微生物接种源与幼苗存活概率相关。(A) 在北部田间实验三年后,存活幼苗上真菌群落的非度量多维尺度 (NMDS) 结果,根据初始微生物接种源的最低温度进行着色。(B) 南方田间实验中存活的幼苗上真菌群落的NMDS结果,根据初始微生物接种源的干旱指数进行着色。(C) 北部田间实验中,每个微生物接种源的幼苗的存活概率与幼苗的平均真菌丰富度的相关性。(D) 在降雨量减少的南部田间地点,每个微生物接种源下幼苗的存活概率与幼苗平均AM真菌多样性的相关性。
微生物介导的气候耐受性与根系相关真菌的多样性有关
即使在田间条件下生长了3年,在两个田间地点和两种菌根类型的幼苗的根系相关真菌群落组成中,初始接种群落仍有统计学意义[perMANOVA,所有的P<0.025;图5A和B,图S3至S6]。这表明,尽管有本地真菌,但至少最初接种的一些真菌类群能够在幼苗根部持续3年以上,这与AM和EM真菌共生体的优先效应相一致。在我们的南部地区,微生物介导的耐热和耐旱性似乎直接或间接地来自于从枝菌根真菌的多样性。这一推断依赖于三个观察结果。首先,来自更干旱地区的初始接种物土壤中AM真菌的多样性更高(LM,P=0.002;图4B和表S12B),即使在田间地头生长三年后,在存活的AM相关幼苗的根部仍然存在这种更高的多样性(PGLMM,P=0.043;图5)。这些模式在 EM 相关幼苗中并不明显。其次,导致 AM 真菌存活的更高多样性的微生物源也往往在降雨减少处理中具有更高的 AM 幼苗存活概率,但在自然环境降雨处理中则不然(GLM,P = 0.015;图 5D)和表 S14B)。第三,这些结果在我们的控制环境实验中得到了证实,在暖干处理中,与AM相关的幼苗的生长与土壤接种物中AM真菌的初始多样性呈正相关(PGLMM,P = 0.001;图4D和表S15)。
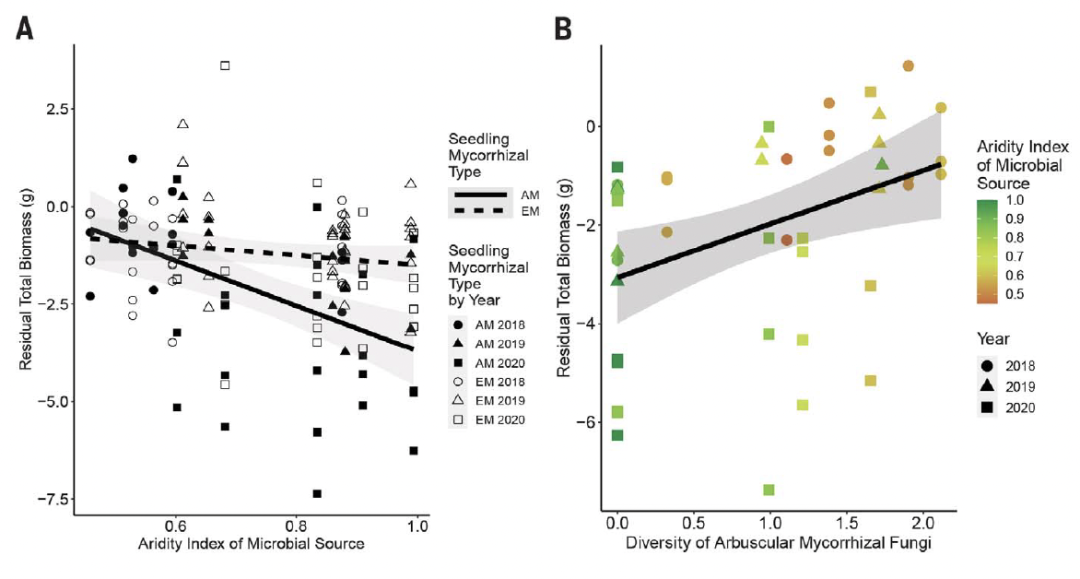
图 5. 控制条件下水分胁迫处理中的幼苗生长反映了 AM 相关幼苗的微生物来源气候。
在控制条件下,研究南部田间的剩余幼苗生物量。残差值解释了幼苗种类和初始大小之间的生长速率差异;正值表示该物种幼苗的生物量大于预期,而负值表示生物量小于预期。(A) 按幼苗菌根类型分开,研究温暖干旱处理中幼苗的残余生物量与初始微生物来源的干旱指数的关系。虚线和开放符号,EM 物种;实线和填充符号,AM 物种。(B) AM 幼苗在温暖干旱条件下的剩余生物量与原始接种物中 AM 真菌多样性的 Shannon-Weaver 指数的关系。原始接种土壤中AM真菌的Shannon-Weaver多样性与干旱呈正相关。
另外,无论菌根类型如何,北部地区的幼苗的耐寒性都会增强。接种了来自温暖地区(较高的最低温度)的微生物源的存活幼苗,在田间地头3年后具有较高的真菌物种丰富度(PGLMM,P=0.027;图5C和表S14A)。与我们南部的田间地点相反,在北部的地点,存活的幼苗上的真菌多样性与环境条件下特定接种源的幼苗的存活概率呈负相关(GLM,P<0.001;图5C和表S16A)。因此,用来自温暖地区的土壤接种的存活幼苗中较高的真菌多样性可能源于接种的真菌群落与田间土生土长的真菌群落的结合。这表明,来自较温暖地区接种的持续存在的适应性差的真菌物种,可能抑制本地真菌的耐寒性。
我们希望微生物诱导的气候耐受性的驱动因素对特定的宿主来说是特异性的。然而,我们的主要结论是,增强植物-宿主耐受性的能力可以通过微生物群落相关的气候来预测。我们的实验涉及一系列的树种和气候变量,表明微生物介导的气候耐受性在温带森林中不是一个罕见的现象。未来的研究应设法确定在生态系统、植物和气候压力下,不考虑具体的微生物机制,是否广泛的发生微生物介导的气候耐受性现象。此外,需要更多的研究来了解微生物群落将如何通过物种的内部选择和物种重新分布来应对不断变化的气候,以及这些变化将对宿主植物的气候耐受性产生什么影响。例如,土壤变暖和干旱实验已经记录了微生物群落组成的变化,但尚不清楚由此产生的新的微生物群落是否会提高植物物种对变暖或干燥的土壤的耐受性。我们的实验表明,在很短的时间内,引入新的微生物类群(通过预先接种的树苗)可以导致更高的植物存活率。
当植物面临越来越大的压力条件时,本土微生物群落对宿主植物的影响可能是有限的,微生物种群可能需要跨空间移动。在这种情况下,我们的实验引入适应性较差的微生物类群会降低物种的存活率。有意地将特定的微生物类群接种到森林中是不可行的。然而,植树是气候战略的一个重要组成部分,人类计划在未来几十年种植数万亿棵树。树木通常是在户外生长后种植幼苗,从而将不同的微生物群落带到种植地点。这些预先接种的树苗代表了根际微生物类群的大规模移动。然而,如果对树木苗圃进行管理,以促进适应特定气候条件的微生物共生体的定植,它们也可能引起微生物介导的气候耐受性。迅速变化的气候已经重塑了生态系统,并导致了物种灭绝。生态系统中,社区成员之间以及跨空间的社区之间的相互联系,有可能放大或降低这些变化。更好地了解微生物的相互作用,作为树木种群在扩展和缩减范围的额外策略,为提高森林的恢复力提供了新途径。
参考文献
Cassandra M. Allsup et al., Shifting microbial communities can enhance tree tolerance to changing climates. Science 380, 835-840 (2023). DOI:10.1126/science.adf2027
- 通讯作者简介 -

威斯康星大学
Richard Lankau
助理教授
Richard Lankau,是威斯康星大学生物系的助理教授,实验室主要研究在自然和农业环境中,植物相关的微生物群落是如何介导个体植物的健康、非生物压力的耐受性和对疾病的敏感性。
猜你喜欢
iMeta简介 高引文章 高颜值绘图imageGP 网络分析iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
系列教程:微生物组入门 Biostar 微生物组 宏基因组
专业技能:学术图表 高分文章 生信宝典 不可或缺的人
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
扩增子分析:图表解读 分析流程 统计绘图
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文,跳转最新文章目录阅读