文章标题:Transcriptomics and metabolomics changes triggered by exogenous 6-benzylaminopurine in relieving epicotyl dormancy of Polygonatum cyrtonema Hua seeds
发表期刊:Frontiers in Plant Science
影响因子:6.627
作者单位:安徽农业大学
百趣提供服务:植物阿趣广靶®代谢组学 研究背景
多花黄精(Polygonatum cyrtonema Hua)是一种药食同源的多年生植物,它是治疗糖尿病和哮喘的重要中药,含有类黄酮和多糖,对抗疲劳和治疗糖尿病有一定作用。但是黄精种子具有“双重休眠”的特征,即胚根休眠和上胚轴休眠;因此,在自然条件下,黄精种子需要经历两个冬天才能顺利出苗,而且繁殖周期非常长,这大大限制了大规模种植。植物激素在植物休眠解除中起着重要作用,6-苄基氨基嘌呤(6-Benzylaminopurine, 6-BA)是一种合成细胞分裂素,通常用于缓解植物的休眠。在本研究中,我们使用转录组学和代谢组学分析了外源6-BA处理后黄精种子的差异。
技术路线
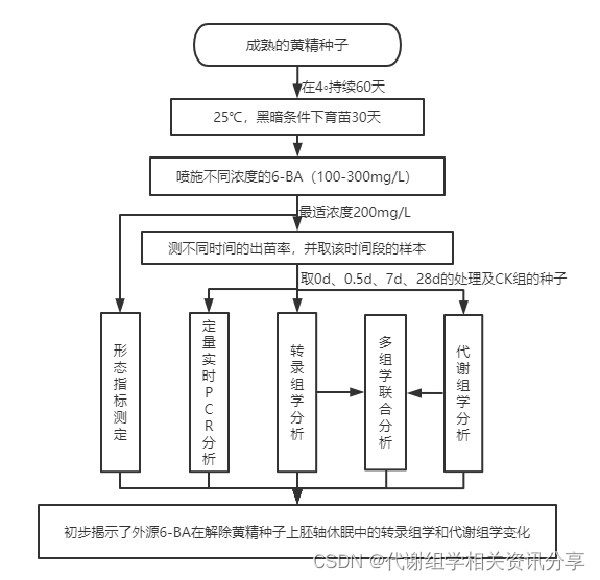
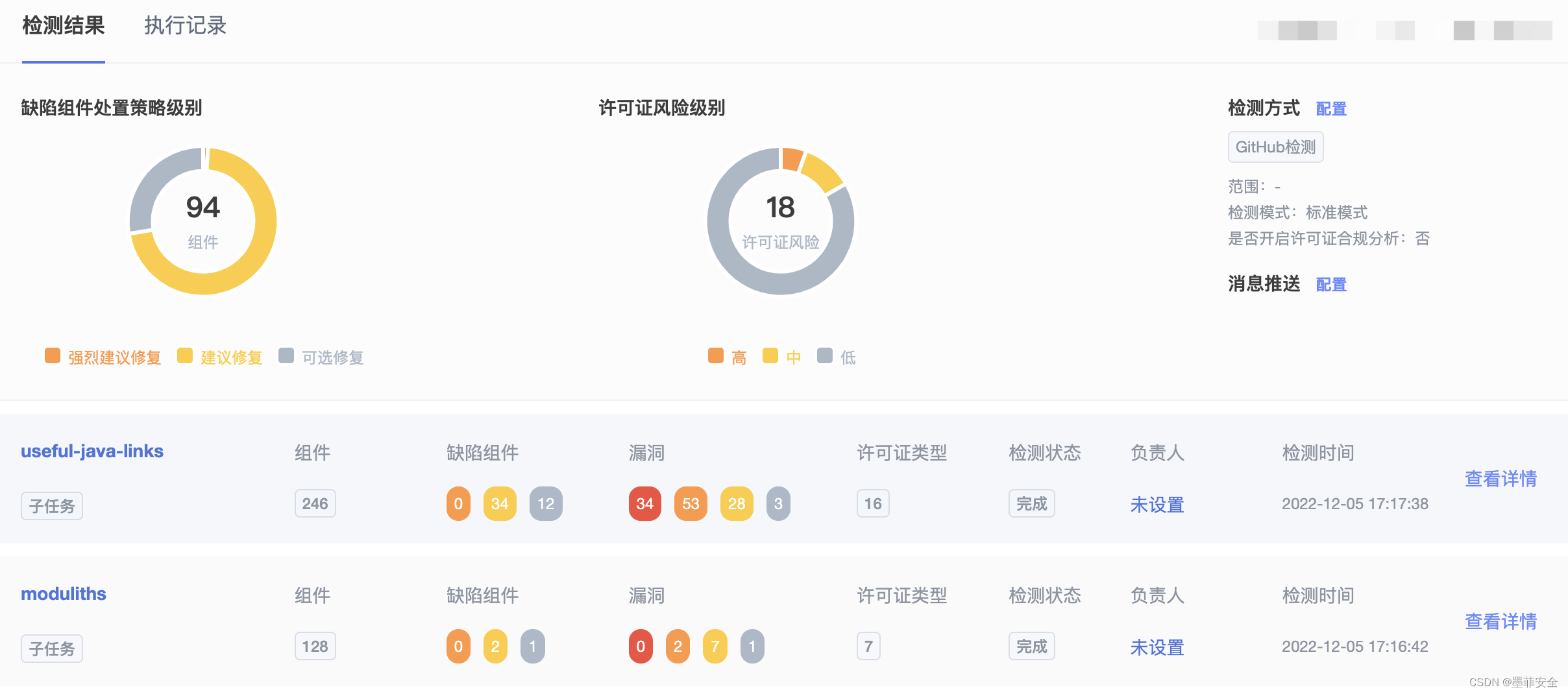
研究结果 1、6-BA处理对黄精种子外观和形态指标的影响
形态学观察表明(图1),黄精种子的发芽率随着培养时间的延长而增加,当发芽率达到60%(30天)时,发芽率增长趋于稳定;当6-BA浓度为100~300mg/L时,效果明显,最佳浓度为200mg/L。处理组在14天时种子相继发芽,28天后出苗率达到60%以上,根部逐渐伸长。但从同一时期的水分对照组来看,进入上胚轴休眠后,上胚轴芽伸长不明显,21天才开始有少量种子发芽,出苗率始终低于10%;但根生长迅速,14天后比处理组长。
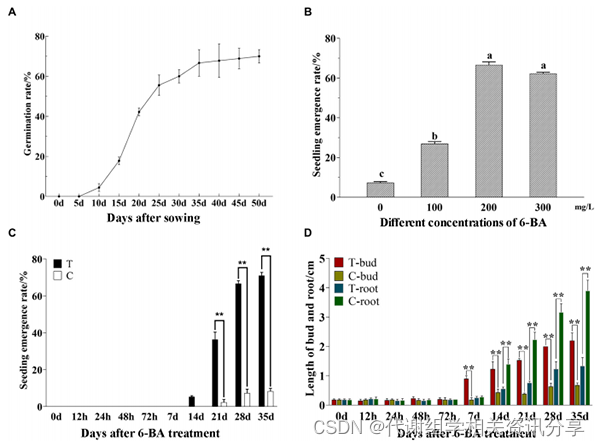
图1. 测定黄精种子种子发芽和出苗形态指标的测定
2、种子萌发过程中黄精的代谢特征
通过植物阿趣广靶®代谢组学(由百趣生物提供)鉴定了21种样品(0天-C0、12小时,7天,28天-C1/T1,C2/T2,C3/T3)代谢物的变化。在样本中共检测到1000种代谢物,包括229种差异代谢物(DEM)。PCA和Venn图分析显示(图2A-D),不同时间段的处理组和对照组之间存在显著差异,其中黄酮类物质存在显著差异。
通过KEGG功能注释分析发现,处理组和对照组在12小时、7天和28天标记了25、33和5条代谢途径(图2E-G)。可以看出,类黄酮生物合成途径、精氨酸和脯氨酸代谢途径在三个阶段显著富集。此外,玉米素生物合成在前两个阶段也显著富集,表明这些代谢途径或代谢产物与休眠解除密切相关。
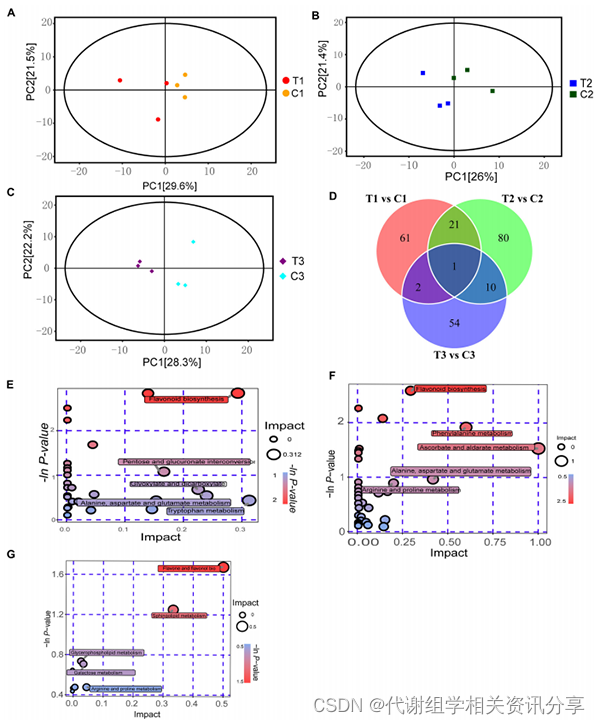
图2. 黄精种子上胚轴休眠解除的代谢组学分析
3、黄精种子不同时期的转录组分析
使用与代谢检测分析相同样品,构建了21个cDNA文库,并进行了高通量RNA-seq分析。通过三个不同阶段的比较分析(图3),作者总共获得了31533个差异表达基因(DEG)。3486、2928和9293个DEG被发现是上调基因,而3213、5221和7392个DEG分别属于下调基因(图3A-B)。有2718、4544和6795个DEG分别在T1与C1、T2与C2和T3与C3的组间比较中差异表达。
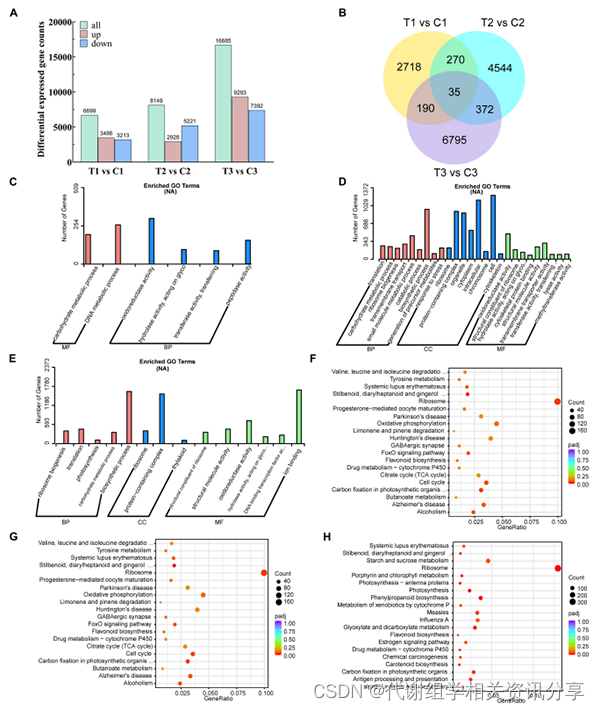
图3. 不同阶段黄精种子的转录组学分析
4、代谢组学和转录组学方法分析了黄精种子上胚轴休眠解除过程
4.1、黄精种子上胚轴休眠解除过程中类黄酮生物合成、精氨酸和脯氨酸代谢以及玉米素生物合成途径中基因和代谢产物的变化(图4)
在6-BA处理解除黄精种子上胚轴休眠的过程中,与同一时期的对照组相比,前7天通过抑制类黄酮生物合成途径的基因和代谢产物来解除对芽分化的抑制,并促进生长素相关基因的表达,从而促进上胚轴芽的伸长。在后期,生长素合成基因的表达受到促进类黄酮相关基因和代谢产物的抑制,这进一步抑制了根系生长,促进了种子向种子出苗的转化。也有报道称类黄酮在种子萌发过程中积累,这表明,黄精种子上胚轴休眠解除和萌发的分子机制不同。
精氨酸和脯氨酸在植物休眠解除中起着重要作用。进一步分析表明,与同一时期的对照组相比,参与精氨酸生物合成的大多数基因和代谢产物在三个阶段中均下调,这可能是因为脯氨酸在这两个阶段没有保持高水平以促进芽伸长。因此,我们认为脯氨酸显著增加了上胚轴芽的伸长,在上胚轴休眠的释放起着重要作用。
玉米素是第一个被发现的植物天然细胞分裂素,我们分析了玉米素生物合成途径的基因和代谢产物的表达模式。总体而言,在前7天,参与玉米素合成途径的基因和代谢物普遍增加,28天后,相关基因和代谢物总体减少,并做了转录组信息验证(图4B–G)。
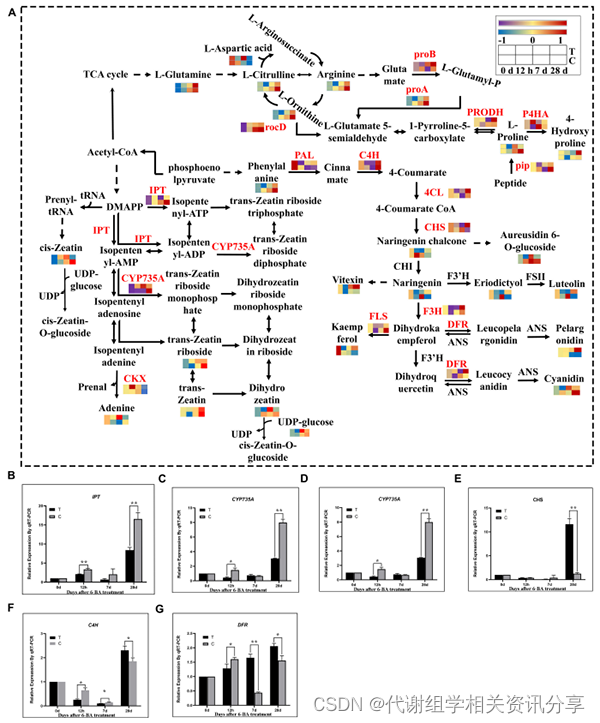
图4. 生物合成途径的转录组学和代谢组学分析。(A)DEG和DEMs在不同阶段参与类黄酮生物合成、精氨酸和脯氨酸代谢以及玉米素生物合成途径。从蓝色到黄色再到红色的颜色梯度,代表基因的低、中、高FPKM。从紫色到黄色再到红色的颜色梯度代表代谢物的低、中、高相对丰度。T:处理组;C:对照组。(B–G)通过qRT-PCR验证转录组中的相关基因。IPT:异戊烯基转移酶基因;CYP735A:细胞分裂素羟化酶基因;CKX:细胞分裂素降解酶基因;CHS:查尔酮合成酶基因;C4H:肉桂酸-4-羟化酶;DFR:二氢黄酮醇-4-还原酶;“*”表示差异显著,“**”表示差异非常显著。
4.2、黄精种子上胚轴休眠解除过程中植物激素途径的基因和代谢产物变化(图5)
植物激素在植物休眠解除过程中起着重要作用,其中较为重要的是吲哚乙酸(IAA)、赤霉素(GA)、脱落酸(ABA)和细胞分裂素(CTK)。
作者对植物激素途径中的DEGs进行计数,最后富集在GA、ABA和IAA代谢途径中。本次研究结果与前人研究结果一致,表明上胚轴芽的伸长在黄精种子休眠解除中的关键作用;属于CTK的玉米素生物合成途径在黄精种子的上胚轴休眠解除中也发挥了重要作用。
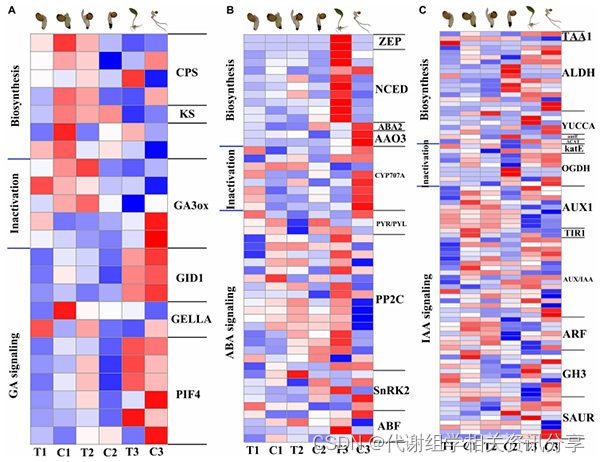
图5. 转录组中GA、ABA和IAA生物合成、失活和信号转导途径相关基因的表达模式。(A)与GA途径相关的基因表达热图。(B)与ABA途径相关的基因表达热图。(C)IAA通路相关基因表达热图。从蓝色到白色再到红色的颜色梯度代表基因的低、中、高表达。Log2(FC)的范围为–1到1。
4.3、类黄酮生物合成、精氨酸和脯氨酸代谢以及玉米素生物合成途径的关联分析(图6)
依据作者先前的结果,我们在T2和C2的三个途径中对DEG和DEMs进行了关联分析(图6)。我们发现精氨酸和脯氨酸代谢途径中的部分代谢物与大多数基因显著相关。此外,在CTK生物合成途径中,参与黄精细胞分裂素脱氢酶基因(PcCKXs)在三个阶段都是活跃的,并与类黄酮、精氨酸和脯氨酸代谢途径代谢产物显著相关,其可能也影响黄精种子的休眠解除。
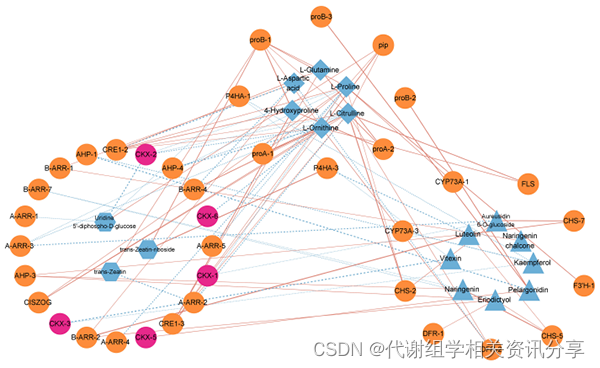
图6. 类黄酮生物合成、精氨酸和脯氨酸代谢以及玉米素生物合成途径的相关分析。注:橙色方框代表基因,蓝色方框代表代谢物,粉色方框代表PcCKX。红线表示正相关,蓝线表示负相关。
4.4、黄精种子上胚轴休眠解除关键时期基因与代谢途径变化模型分析
作者依据多组学分析结果构建模型(图7),得到外源6-BA处理促进GA生物合成和信号转导途径,并诱导黄精种子上胚轴休眠的释放。同时,外源6-BA处理可通过抑制PcCKXs的表达,增加上胚轴芽伸长过程中内源性细胞分裂激素的含量。未来的研究将集中于这些关键因素在黄精种子休眠解除中的功能作用。
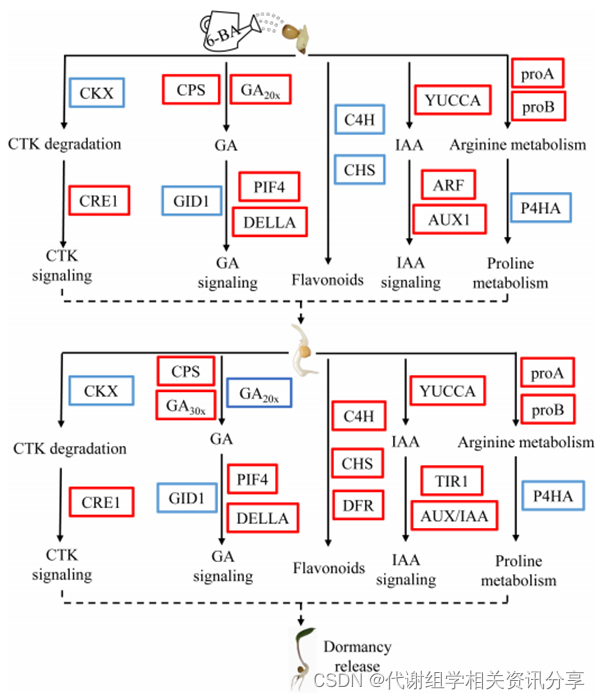
图7. 黄精种子上胚轴休眠解除关键时期代谢途径变化示意图模型。 结论
已有研究证实外源GA3可以缓解种子休眠,但不能缓解黄精种子的上胚轴休眠。在本研究中,作者探讨了外源6-BA处理对黄精种子上胚轴休眠解除的分子机制。结果表明,外源6BA处理能显著延长上胚轴的芽长,提高黄精种子的出苗率。在黄精种子上胚轴休眠解除过程中,处理组的玉米素、脯氨酸、生长素和赤霉素含量较高,而类黄酮和精氨酸含量较低。PcCKXs与类黄酮生物合成代谢物以及精氨酸和脯氨酸代谢物具有显著相关性,这些结果为外源6-BA解除黄精种子上胚轴休眠的分子机制提供了新的见解。