湖南农业大学科研团队揭示水稻“生长—防御”平衡调控机制!
文章标题:Rice cellulose synthase-like protein OsCSLD4 coordinates the trade-off between plant growth and defense
发表期刊:Frontiers in Plant Science
影响因子:6.627
作者单位:湖南农业大学
百趣生物提供服务:植物阿趣广靶®代谢组学
1.代谢组学分享-研究背景
通常植物防御能力的提升伴随着生长和繁殖成本的降低,这是一种在不利条件下微妙的适应性机制,被称为生长-防御权衡机制。纤维素合成酶类蛋白(cellulose synthase-like D4, OsCSLD4)通过调节细胞壁多糖的合成,在水稻的植物结构中发挥重要作用。代谢组学分享,近期研究表明,OsCSLD4通过介导脱落酸的生物合成参与水稻的盐胁迫反应。然而,OsCSLD4在水稻对生物胁迫反应中的作用机制尚未可知。在本研究中,利用合江19号水稻突变体中的叶片(dwarf and narrowed leaf in Hejiang 19 mutant, dnl19)来确定OsCSLD4在水稻生长和防御之间发生的作用,揭示了OsCSLD4在调控水稻生长和防御之间的平衡中的分子机理。
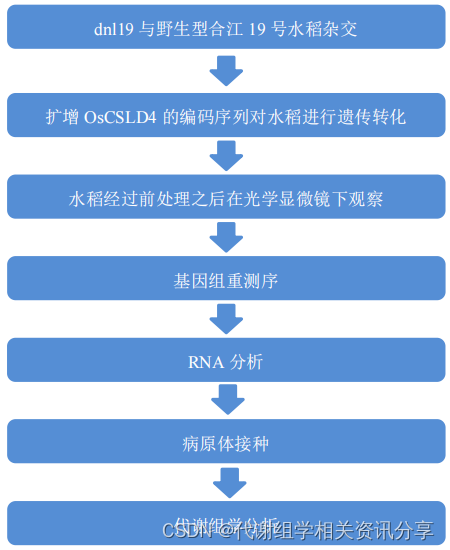
2.代谢组学分享-主要实验
3.代谢组学分享-研究结果 水稻dnl19突变体的多态性表型
本研究检测了dnl19突变体的旗叶长度和宽度、圆锥花序长度和节间伸长率。结果表明,在整个发育期,dnl19突变型水稻比野生型(wild type,WT)矮(图1)。代谢组学分享,另外,与WT相比,dnl19突变体植株的叶宽、穗长、结实率以及粒径受到影响,粒宽和千粒重显著下降。总之,dnl19突变体植株表型整体上有很大变化。
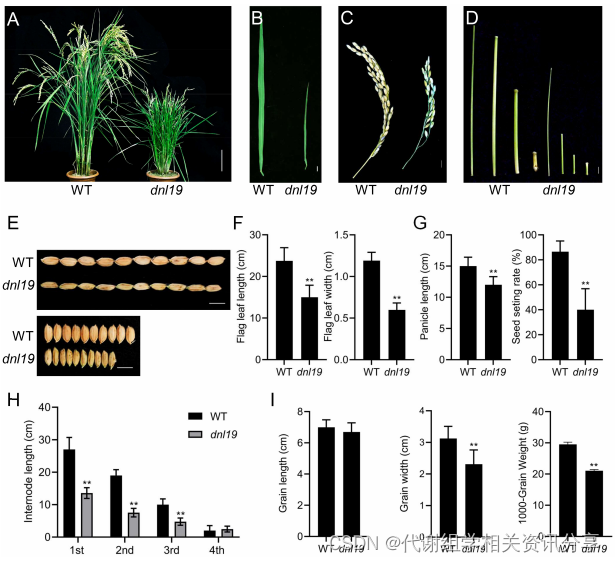
图1. dnl19突变体的表征 dnl19中叶脉图案和节间细胞大小的改变
通过对叶片等部位进行组织切片观察,发现突变株小叶脉减少进而引起窄叶表型变化,而透明细胞的显著减小引起中叶脉厚度的减小。代谢组学分享,并且,dnl19突变株在叶脉区具有明显比野生型更小的泡状细胞,这表明dnl19叶片容易卷曲。对第二节间纵切面的检测结果表明,细胞数量减少导致第二节间长度缩短。
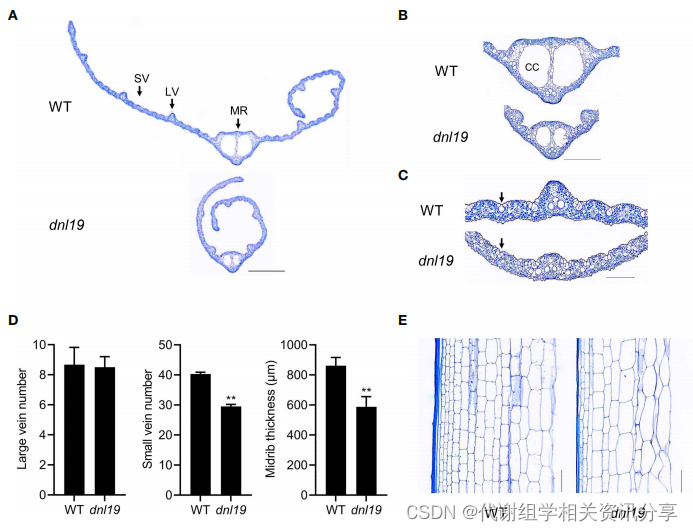
图2. dnl19突变体对叶片和茎秆结构的影响
dnl19突变体T-DNA插入的基因组侧翼序列的分离
通过高效热不对称交错PCR技术对dnl19突变体的T-DNA插入位点进行鉴定。代谢组学分享,结果显示,T-DNA插到了CHR702的内含子区域。研究证明,T-DNA插入突变体以及CHR702敲除未引起任何形态上的差异变化。此外,部分未经转基因的dnl19也具有矮小的形态。因此,推测CHR702不是dnl19突变体的致病基因。
通过MutMap分析鉴定dnl19突变体的候选区域和致病SNPs
通过分别对合江19号、dnl19突变株、正常组和矮化组样品进行测序深度为30×的全基因组重排分析。代谢组学分享,结果表明,在候选区域的单核苷酸多态性(SNP)中,大多数 SNP指数差异(△SNP指数)大于0.6的基因位于非编码区。位于22,606,187号核苷酸位置的SNP位于基因OsCSLD4的第一外显子中。先前的研究表明该基因具有调节细胞壁多糖合成的功能。在密码子302处有一个C碱基缺失引起移码,并且过早引入了一个终止密码子。测序分析证实突变位点和C碱基缺失可能是dnl19突变体表型异常的原因。
为了挽救dnl19突变体的各种发育缺陷,作者在玉米泛素启动子的控制下在dnl19 植物中表达了OsCSLD4。代谢组学分享,将该基因成功引入突变体dnl19后,dnl19的表型被完全抑制。至于植株高度和叶片的形态特征,各植株间没有差异。
表1. 染色体基因组中△SNP指数>0.6的SNP的候选基因
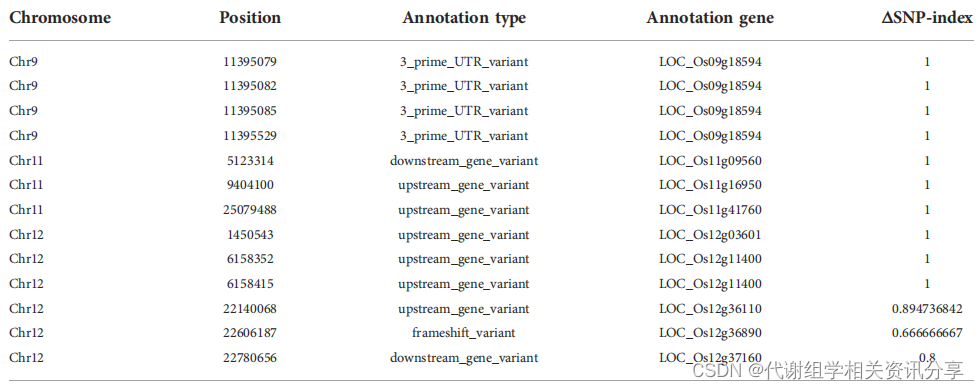
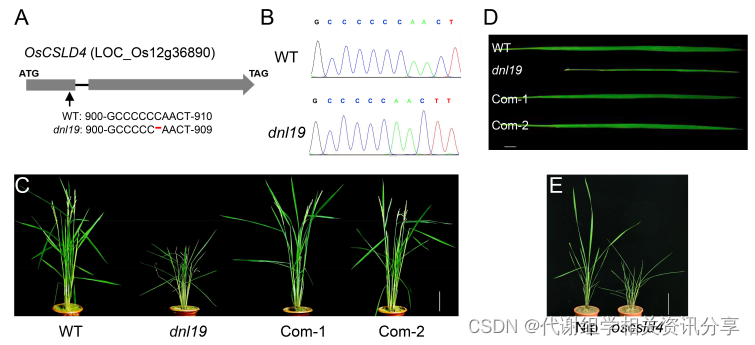
图3. dnl19突变体的致病基因(OsCSLD4) 通过RNA测序对dnl19突变体进行转录组分析
在植物中,生长发育受损通常伴随着防御能力增强。为明确OsCSLD4是否参与水稻防御反应,本文作者利用RNA-seq技术对抽穗期dnl19和WT旗叶进行转录组检测,并结合qPCR技术检测dnl19和WT在幼苗期和分蘖期的一些防御反应基因的表达水平。代谢组学分享,dnl19的PR基因(致病相关基因)在14日龄时显著上调。PR基因中的其他基因在幼苗期和分蘖期都有不同程度地增加。这些结果都表明dnl19中 OsCSLD4 的破坏激活了防御反应基因的表达。

图4. dnl19突变体和野生型的转录组分析
dnl19突变体对稻瘟病和水稻黄单胞菌的抗性发生改变
作者在苗期通过打孔接种对dnl19进行抗稻瘟病的能力评估。结果表明,接种稻瘟病菌110-2菌株后,dnl19叶片的病斑明显减少。对每个病斑的相对真菌生物量进行研究,突变体叶片中生长的真菌较少。作者还评估了dnl19突变体对白叶枯病的抗性。通过检测分蘖期接种PXO99的日本晴叶片中OsCSLD4的表达。代谢组学分享,结果显示,处理48小时后的OsCSLD4表达显著增加。然后通过顶切方法将PXO99接种到dnl19突变体的旗叶上。接种2周后,dnl19的病斑长度显著短于WT。以上结果表明OsCSLD4基因功能的缺失增强了水稻的抗病性。
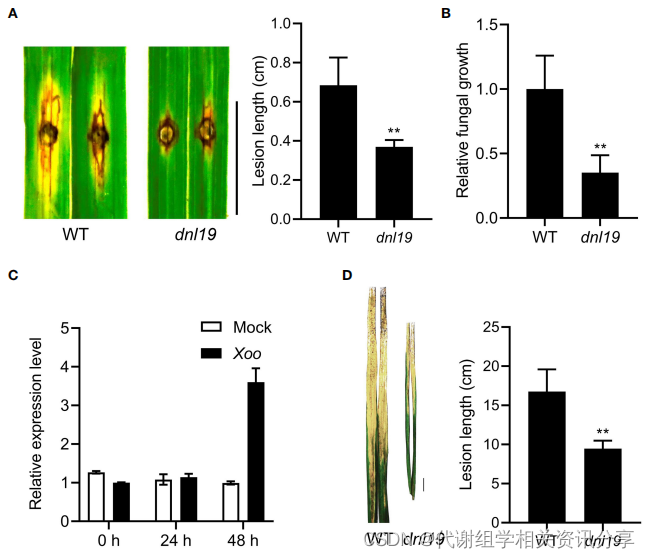
图5. dnl19突变体和野生型对稻瘟病和水稻黄单胞菌的抗性表征
dnl19突变体的代谢分析
为了了解OsCSLD4突变的代谢反应,作者应用广泛靶向代谢组学来研究dnl19突变体的变化情况(图6)。在dnl19突变体中共鉴定出69种不同差异代谢物。代谢组学分享,其中,上调的差异代谢物有26种,包括L-缬氨酸、L-天冬酰胺、L-组氨酸、L-丙氨酸、龙胆酸和其他重要代谢产物;下调的差异代谢物有43种,包括L-天冬氨酸、苹果酸、6-磷酸葡萄糖酸、6-磷酸葡萄糖、1-磷酸半乳糖、葡萄糖酸、D-天冬氨酸等(图6C)。分析结果表明,dnl19中OsCSLD4的破坏导致L-缬氨酸、L-天冬酰胺、L-组氨酸、L-丙氨酸、龙胆酸的显著增加,但L-天冬氨酸、苹果酸、6-磷酸葡萄糖酸、6-磷酸葡萄糖、1-磷酸半乳糖、葡萄糖酸、D-天冬氨酸的显著减少。
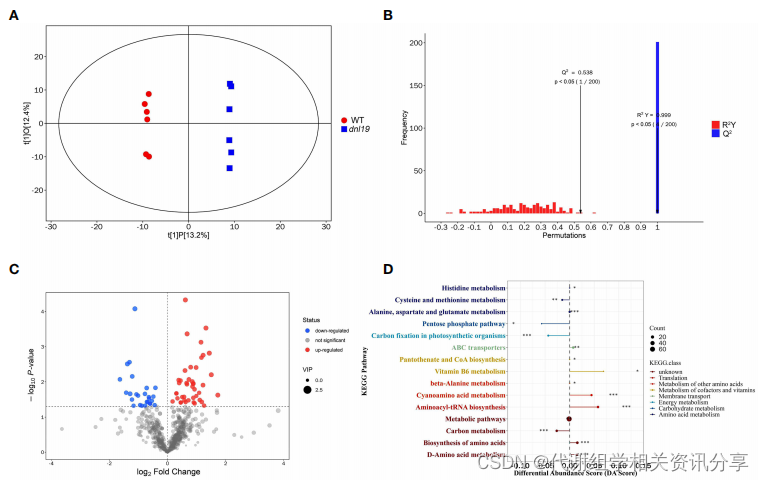
图6. dnl19突变体的广泛靶向代谢组学分析
4.代谢组学分享-结论
本研究利用dnl19进行基因组学和代谢组学的研究分析,探讨了OsCSLD4在平衡水稻生长和防御之间的权衡中的重要性。代谢组学分享,一般来说,细胞壁多糖合成酶在水稻生长和非生物胁迫中具有关键作用,在病原体感染期间,特定低聚糖作为主要的植物源性损伤相关模式分子(damageassociated molecular pattern, DAMP)产生,并可以激活植物免疫反应。然而,尚不清楚细胞壁生物合成缺陷的dnl19或其他OsCSLD4缺失突变体是否也会释放或提供寡糖信号分子的前体,以组成性地触发植物防御反应,还需要进一步研究。