
文章内容导读
叶绿体和线粒体是唯一拥有内共生起源基因组的细胞器。近年来,测序技术的进步促进了大量植物细胞器基因组的发表,并揭示了它们之间极大的进化轨迹差异。本综述量化了在植物进化树中分布广泛的已测序植物细胞器基因组的数量和分布情况。
文章内容亮点
● 技术的进步导致植物细胞器序列数量大幅增加,但取样仍然不均衡;
●大多数陆生植物线粒体基因组由于替代结构配置和转移序列的干扰而难以准确组装;
● 泛细胞器基因组提供了新的进化见解,并为研究谱系分歧期间基因功能的变化提供了编辑目标;
● 细胞器基因组编辑——如线粒体靶向转录激活样效应物核酸酶(TALENs)和双链DNA特异性胞嘧啶脱氨酶(DddA)源自胞嘧啶碱基编辑器 —— 使开放阅读框(ORFs)和非编码区的功能验证成为可能;
● 叶绿体转化被用于一系列修饰,例如提高光合效率、疫苗生物合成等,而线粒体转化由于技术挑战而远远落后。
主要内容
01、植物细胞器基因组研究进展
与大型的真核基因组相比,大多数叶绿体和植物线粒体基因组要小得多。虽然已经测序的完整叶绿体基因组的数量接近13,000个,但完全测序的植物线粒体基因组只有673个,且且两者均测序的物种只有285种。叶绿体基因组数量激增主要得益于测序技术进步,但线粒体基因组完成度仍滞后(图1A、B)。短读长测序方法在线粒体叶绿体基因组中的表现不佳,主要受重复序列和转移片段影响。
截止2023年9月在NCBI上已经发表了近13,000个叶绿体基因组(plastomes)(图1),其中绝大多数来自被子植物,尤其是双子叶植物(图1A)。
然而,相对于测序的植物线粒体基因组,被子植物的百分比显著降低。由于其结构复杂、组装难度大,已测序的线粒体基因组数量并不能完全反映生物多样性。目前,经济重要性和谱系丰富的物种仍然是关注的重点,而第三代测序技术的应用也为更多的线粒体基因组测序提供了可能性。目前,针对植物线粒体基因组的特定组装工具及评估体系仍需开发。
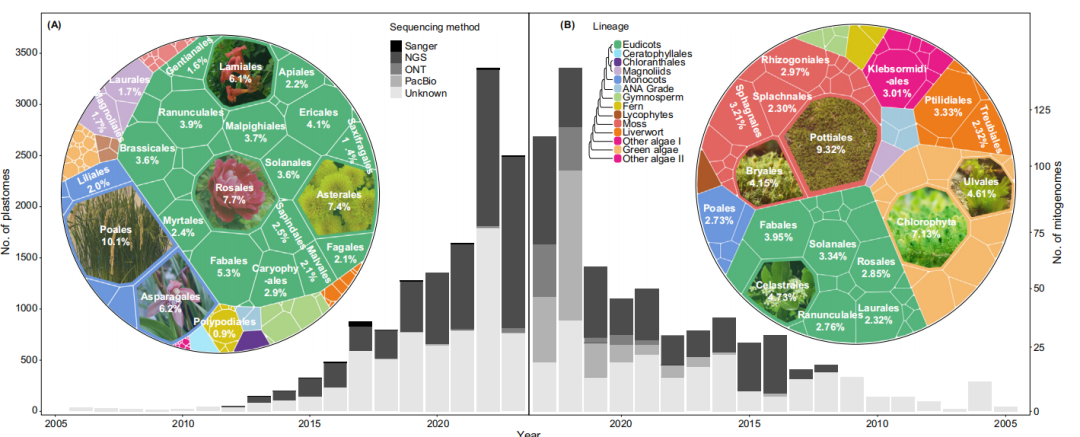
图1 已发表的植物细胞器基因组的测序现状
02、植物细胞器基因组的演化与功能差异
基因内容不同
在内共生演化过程中,两种细胞器的基因内容发生显著衰减。但光合作用和呼吸电子传递链等机制基本完整。大多数叶绿体包含约100个基因,包括编码蛋白基因(PCG)、rRNA和tRNA。植物线粒体基因组包含的功能基因数量少但总体长度大。目前不同物种的线粒体基因数之间存在较大差异,从14个(寄生植物Viscum minimum)到50个(苔类Marchantia polymorpha)不等。基因丢失和tRNA的含量变异也普遍存在。线粒体基因组与叶绿体基因组之间的基因内容差异取决于功能性基因向细胞核基因组转移的程度以及在进化过程中是否被核基因取代。
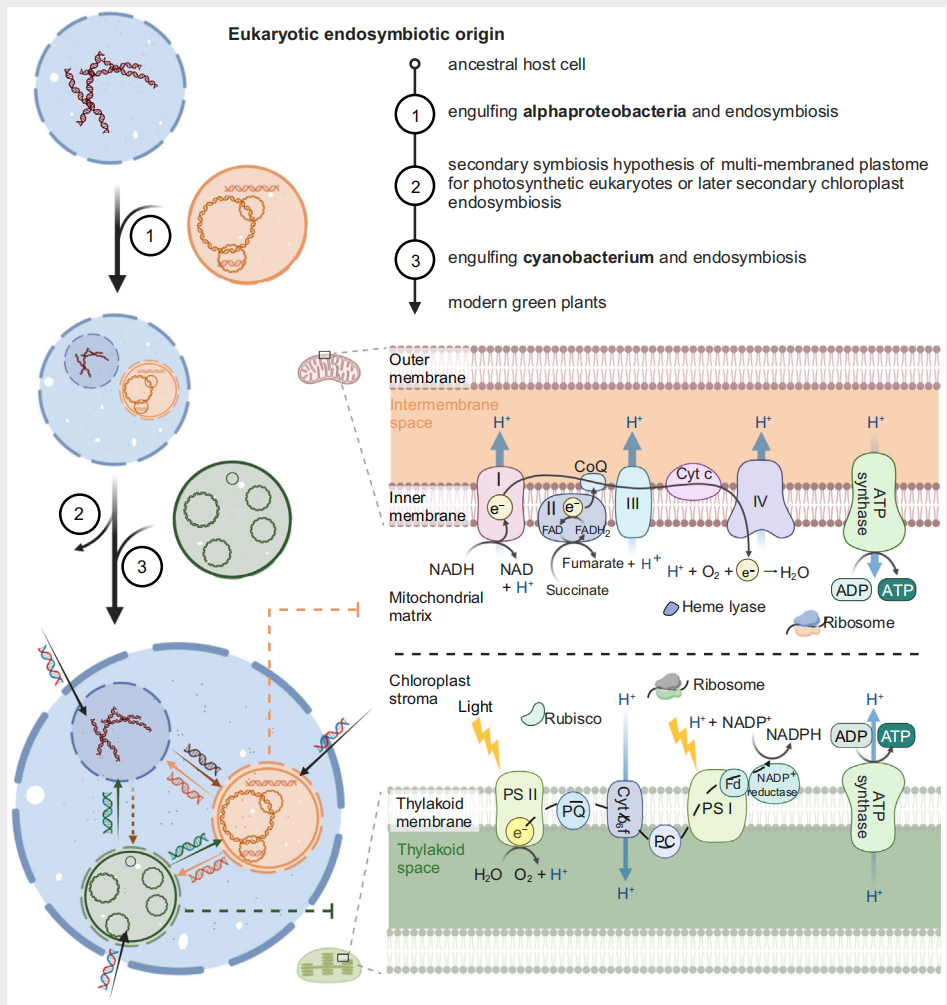
图2 细胞器基因组的内共生起源
突变率不同
植物线粒体基因组结构迅速演化而序列演化缓慢。核基因组演化最快,叶绿体次之,线粒体则最慢。叶绿体突变率在不同谱系中变化不大,但线粒体突变率差异显著。远缘植物谱系中存在异常速率变异(例如,Pelargonium、Plantago和Silene)。机制塑造细胞器基因组中替代速率的机制主要包括;选择压力、突变负担和局部分化超突变。这些机制涉及多个负责目标细胞器复制、修复和重组(RRR)的核调节基因。
结构变化的差异
几乎所有叶绿体基因组都具有高度保守的四部分环形结构,由一个大的单拷贝(LSC)区和一个小的单拷贝(SSC)区组成,两部分之间隔以一对IR区。除此之外,叶绿体基因组仍包含循环、线性和多分支等多种形式;循环形式仅占一小部分。在典型的叶绿体基因组中,两种构型共存,并通过IR进行互换(图2C)。但也存在一些特殊现象,例如在松属中IR极度缩短、在Papilionideae中整个IR丢失、在Selaginellaceae的一些物种中直接重复(DR)以及除固定形式外的动态结构网络(图2D)。
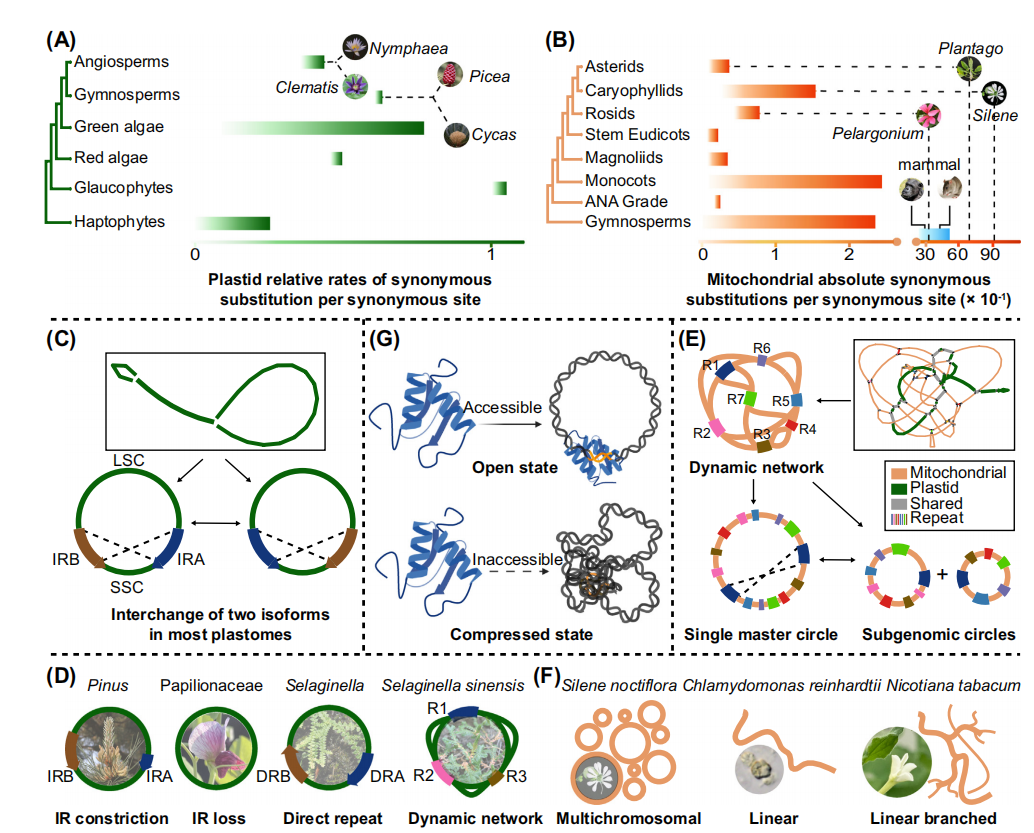
图3 植物细胞器基因组的突变和结构变异
03、转移事件塑造了植物细胞器基因组的进化
植物细胞器基因组中的特有现象是水平基因转移(HGT)和内源性基因转移(IGT)。涉及细胞器的HGT事件的报告很少,主要集中在寄生植物线粒体中。通过寄生虫与宿主或共生体间物理接触发生,是生物进化的重要驱动力。
植物细胞内的IGT是持续动态发生的,能增加植物基因组复杂性,但其转移模式随机,缺乏统一规律。随着基因组数据的增加,研究IGT在更广泛分类学水平上的规律性和机制成为可能,这对于理解其在塑造植物基因多样性中的作用至关重要。
04、细胞器基因组编辑的进步与局限性
线粒体基因组中的序列突变和结构变异会影响基因功能,尤其是在频繁重组导致与细胞质雄性不育(CMS)和其他未知功能有关的嵌合ORF的植物叶绿体基因组中。目前,几种编辑技术显示出巨大的潜力用于叶绿体基因组编辑(图3),涉及到了多种基因编辑工具的原理、特点和适用范围,如MitoTALENs、MitoDdCBEs、CRISPR/Cas9等,不同类型的基因编辑在核基因组和质体基因组中的应用情况是有所不同的(具体内容可见文章对该章节的详细描述)。
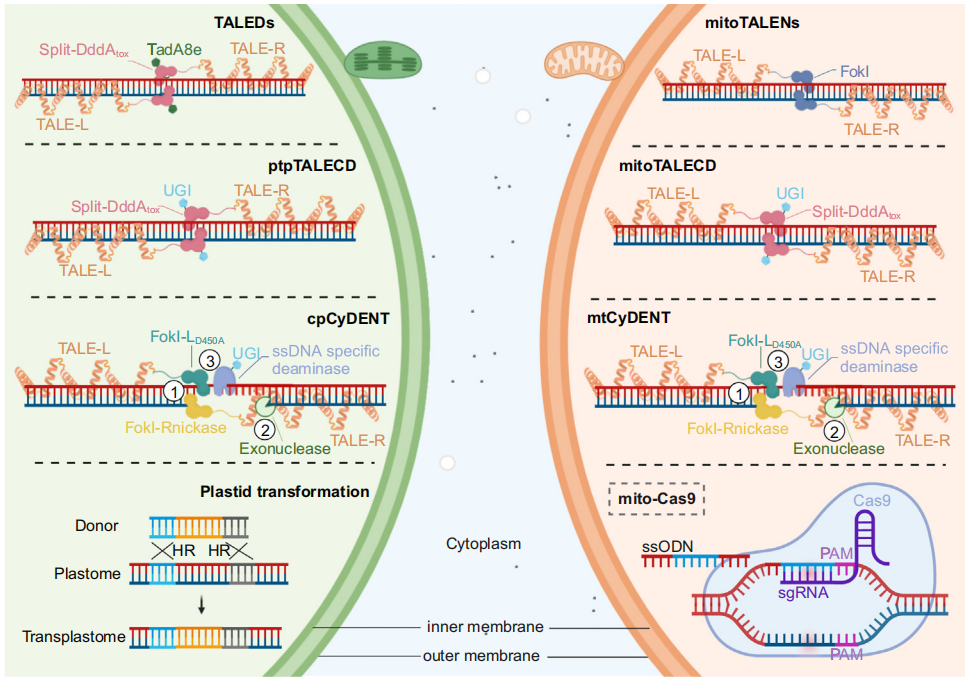
图4 细胞器基因组编辑技术示意图
05、植物细胞器基因组的未来研究与展望
应不断开展大量与细胞器相关的研究,以提高研究人员对植物细胞器的理解。长读长测序技术应该与三维基因组学和电子显微镜相结合,以提高解决替代配置准确性的能力。泛3D细胞器基因组是研究组织、发育阶段、个体和物种之间微进化的重要下一步。还应更详细地研究许多ORF和被忽视的非编码区域,以了解它们在植物细胞器功能和进化中的作用。在细胞器基因组编辑方面有许多独特的优势。优化现有工具并开发新的具体编辑技术同样重要,以提高编辑效率,特别是直接递送到细胞器的技术,这是研究植物细胞器基因组所必需的。
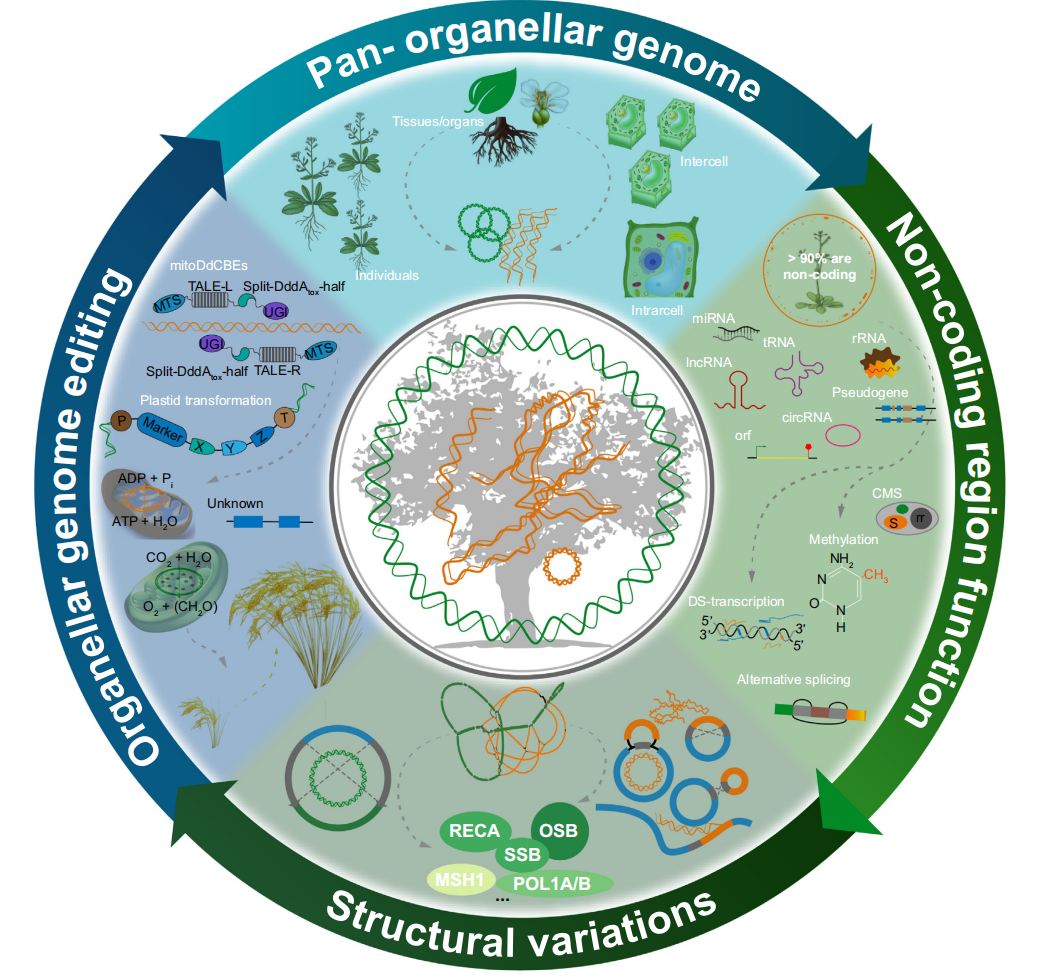
图5 泛细胞器基因组(上图)的目的是重新评估个体、组织、发育阶段、细胞间和分子内群体之间的异质性和遗传性
凌恩研发团队在细胞器基因组研究方面积累了丰富经验,并建立了成熟的研究技术流程。在解决上述难点基础之上,成功协助了许多研究人员实现科研目标,并发表了大量高水平的学术论文,欢迎来电咨询。
凌恩细胞器研究技术优势:
1、提供专业mtDNA&cpDNA富集抽提服务,自主研发的细胞器提取技术,高效实现高质量细胞器基因组富集;
2、告别在线注释平台的偏差,满足NCBI数据库上传要求,高质量结果交付,助力高水平研究;

参考文献
Plant organellar genomes: much done, much more to do?Trends in Plant Science, 2024.



















