文献介绍

「文献题目」 Integrating barcoded neuroanatomy with spatial transcriptional profiling enables identification of gene correlates of projections
「研究团队」 Anthony M. Zador(美国冷泉港实验室)
「发表时间」 2021-05-10
「发表期刊」 Nature Neuroscience
「影响因子」 24.8
「DOI」 10.1038/s41593-021-00842-4
摘要
功能环路由具有不同轴突投射和基因表达的神经元组成。理解投射神经元的分子特征需要在细胞分辨率上同时对基因表达和同一细胞中多个靶点的投射进行高通量询问,这使用目前的技术很难实现。在这里,作者介绍了 BARseq2,一种通过原位测序同时绘制投射并检测多个基因表达的技术。作者确定了 29,933 个细胞中钙粘蛋白和细胞类型标记的表达,以及小鼠 motor cortex (运动皮层) 和 auditory cortex (听觉皮层) 中 3,164 个细胞的投射。在 1,349 个神经元中的相关基因表达和投射显示了两个皮层区域同源投射的钙粘蛋白特征。这些钙粘蛋白在转录组分类的多个分支中富集。通过在单个神经元中高通量将多基因表达和投射到多个靶标相关联,BARseq2 为揭示神经元环路的分子逻辑提供了一种潜在的方法。
研究结果
为了研究钙粘蛋白表达与不同投射的关系,作者开发了 BARseq2,将高通量投射绘制与使用原位测序的基因表达多重检测相结合(Fig. 1b,c)。BARseq2 是基于 BARseq (Fig. 1c),通过 RNA 条形码的原位测序,实现了高通量的投射绘制。使用 BARseq 观察到的投射模式与在多个环路中使用传统神经解剖技术获得的投射模式一致,但它可以实现比最先进的单细胞追踪技术高出至少 2 到 3 个数量级的通量。可能的技术问题,包括对通道纤维与轴突末端的区分、灵敏度、神经元的双标记和条形码的退化,以前已经讨论了这里将不再详细讨论。将条形码单细胞投射绘图与内源性 mRNAs 的原位检测相结合,利用 BARseq 在通量方面的独特优势,有效地同时询问神经元基因表达和长程投射。
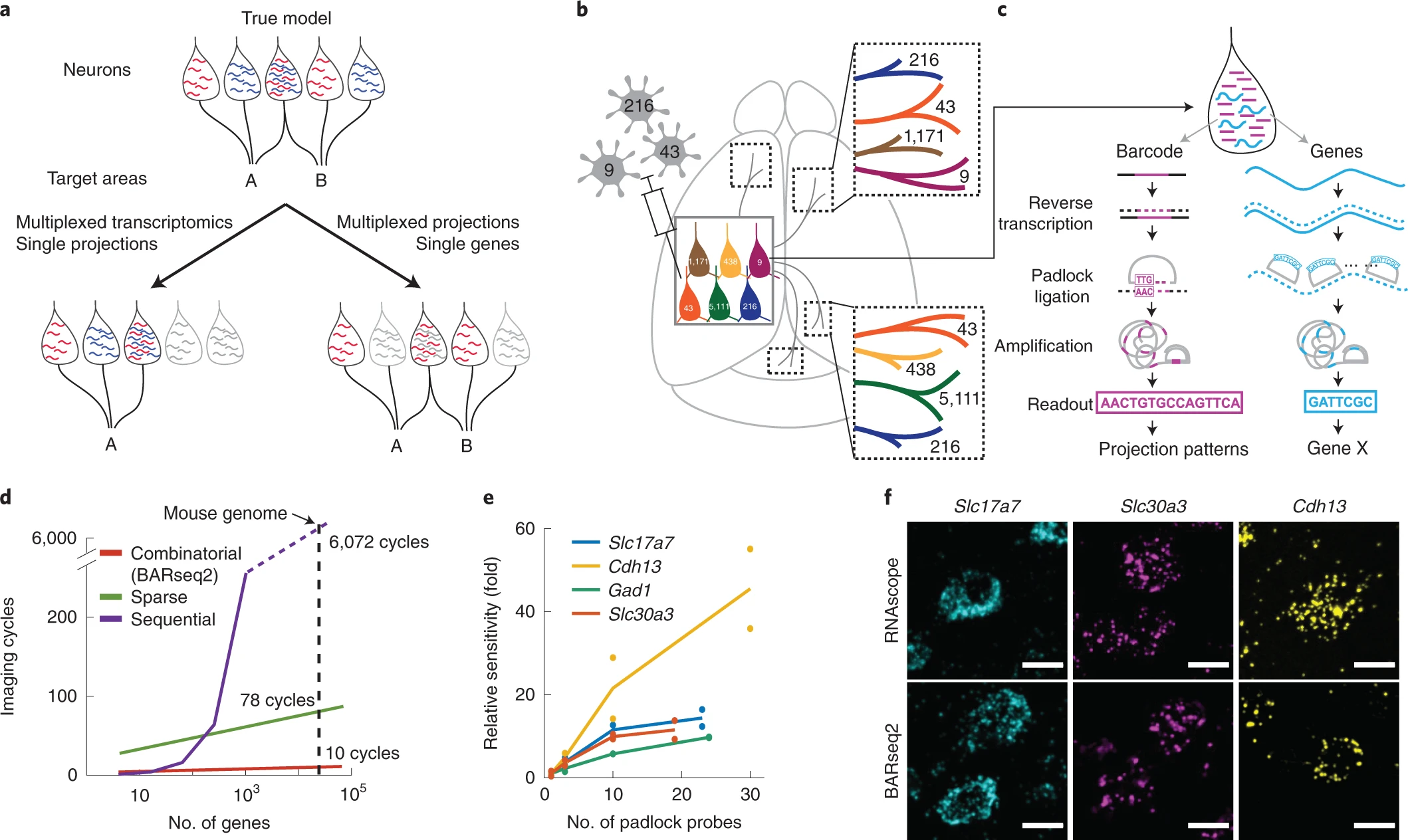
a.一个卡通示例模型,投射和基因表达之间的关系只能通过对投射和基因表达的多次询问来正确地推断。在这个模型中,表达两个基因的神经元同时靶向到 A 和 B,而只表达两个基因之一的神经元随机靶向到 A 或 B 中的一种。将单个神经元多基因表达与只有一个投射靶点的数据相结合的方法,将会得到这样的结论:这三种基因表达模式都投射到靶标 A,因此无法检测到基因表达和投射之间潜在的“真实”关系。同样地,将多个单神经元投射与单个基因的数据相结合的方法,也将无法检测到基因表达和投射之间的任何关系。
b-c.BARseq2 相关投射和基因表达在细胞分辨率上 (b)。在 BARseq2 中,神经元用随机的 RNA 序列进行条形编码来实现投射绘制,基因也在相同的条形编码神经元中被测序。RNA 条形码和基因被扩增和读取使用不同的策略(c)。
d.使用组合编码的理论成像周期 (BARseq2),Eng 等人所使用的四通道顺序编码或四通道稀疏编码的理论成像周期。成像周期假设 BARseq2 为三个额外周期,一轮的稀疏编码,没有额外周期的纠错。
e.BARseq2 在使用每个基因不同数量的锁式探针检测指示基因时的相对灵敏度的平均和单个数据点。灵敏度被归一化为使用一个探针每个基因。每个基因 2 个切片 (n=2)。
f.与 RNAscope 相比,使用 e 中显示的最大探针数对指定基因进行 BARseq2 检测的代表性图像。比例尺,10 µm。
为了使用 BARseq2 检测基因表达,作者使用了一种基于无间隙填充锁式探针(non-gap-filled padlock probe-based)的方法来放大目标内源性 mRNAs (Fig.1c)。消除间隙填充是读取极其不同的条形码序列所必需的,它增加了内源性基因检测的灵敏度。在这种方法中,通过使用原位 Illumina 测序化学方法测序基因识别指数 (gene identification index, GII) 来读出靶点的身份。由于 GII 是一个核苷酸条码序列,它唯一地编码给定基因的身份,因此多重编码能力以指数级增长为 4N,其中 N 是测序循环的数量。这种组合编码通过序列读出,从而允许使用少量几个周期的成像来同时检测大量基因 (Fig. 1d)。尽管测序读数提供了许多优势,但必要时 BARseq2 也与基于杂交的读数兼容。内源性基因的原位测序和条形码测序的间隙填充方法相结合,允许许多基因与使用 BARseq2 的投射同时被检测。
作者首先证明,通过优化靶向原位测序,BARseq2 有足够的灵敏度来检测内源性 mRNAs。接下来,作者将内源性 mRNA 的原位测序与 RNA 条形码的原位测序结合起来,将钙粘蛋白的表达与细胞分辨率下的投射模式联系起来。然后,作者通过证明验证了 BARseq2,它可以用于概括转录组神经元亚类型的投射模式,并识别在主要投射类中不同表达的钙粘蛋白。最后,我们确定了小鼠听觉皮层和运动皮层之间共享的钙粘蛋白,它们与两个皮层区域脑内 (IT) 神经元的同源投射相关。
1. BARseq2 稳健地检测到内源性 mRNAs
为了使用 BARseq2 充分检测基因,作者试图提高检测灵敏度。在大多数原位杂交 (in situ hybridization, ISH) 方法中,通过对每个靶点 mRNA 使用多个探针可以提高灵敏度。作者认为,增加每个基因的锁式探针的数量可能同样会提高 BARseq2 的灵敏度。事实上,作者观察到,与使用单个探针相比,使用多个探针的灵敏度增加了 46 倍 (Fig. 1e and Methods)。结合其他技术进行优化(Extended Data Fig. 1a,b),作者将 BARseq2 的灵敏度提高到 RNAScope (一种灵敏的和商业上可行的 FISH 方法) 的 60% (Fig. 1f, Extended Data Fig. 1c,d and Methods)。作者进一步优化了原位测序,以在许多测序周期中稳健地读出单个细胞的 GIIs (Extended Data Fig. 1e–j and Methods)。这些优化允许 BARseq2 实现对 mRNA 进行足够灵敏、快速和稳定的检测。
2. BARseq2 允许原位检测多个 mRNAs
为了评估使用 BARseq2 多重原位检测钙粘蛋白,作者检查了 20 个钙粘蛋白的表达,以及 3 个(听觉皮层)或 45 个(运动皮层)细胞类型标记 (Fig. 2a–c)。作者选择关注钙粘蛋白,因为它们在皮层发育中的作用已知,包括投射特异性,以及它们在由多个属性定义的基本细胞类型之间的差异表达。这些钙粘蛋白类包括大多数经典听觉皮质和运动皮质和非聚集类原钙质素。作者成功地解决并解码了 2 张小鼠听觉皮层切片 (1.7 mm2 × 10 µm per slice) 的 419,724 个 rolonies,以及 4 个初级运动皮层切片 (2.8 mm2 × 10 µm per slice) 的 1,445,648 个 rolonies。作者在听觉皮层中恢复了 20 个 rolonies 和运动皮层中 115 个 rolonies,与实验中没有使用的两个 GII 匹配,rolony 解码的估计错误率分别为 0.1% 和 0.2%。
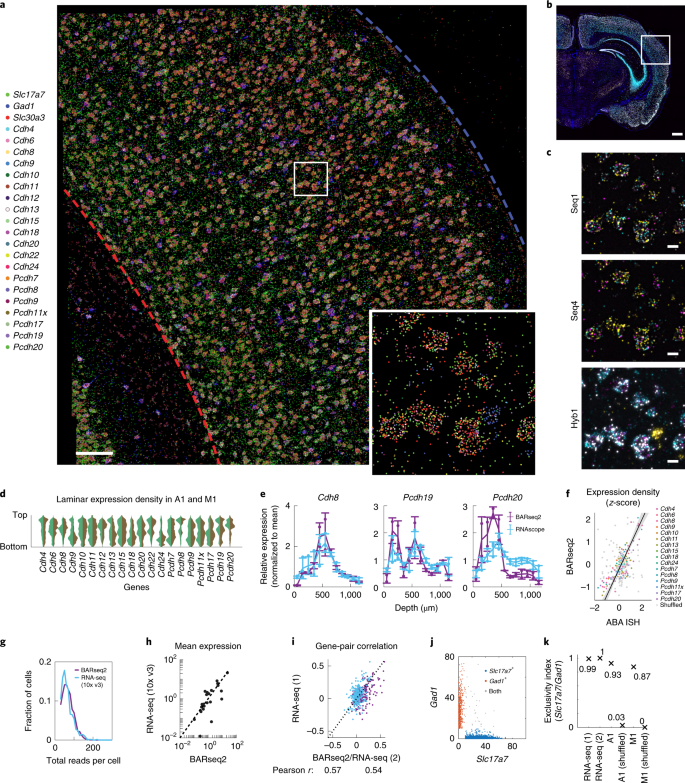
a.听觉皮层中 rolonies 的代表性图像(来自两个切片的测序)。大脑皮层的顶部和底部分别由蓝色和红色的虚线表示。比例尺,100µm。嵌入部分显示方框区域的放大视图。
b.杂交循环的低放大图像,显示在 a 中成像的区域的位置。比例尺,100µm。
c.在 a 中的方框区域的指示的测序轮数和杂交轮数的代表性图像。比例尺,10µm。
d.小提琴图显示了钙粘蛋白表达在神经元胞体中的层状分布。听觉皮层(绿色)和运动皮层(棕色)。
e.由 BARseq2 或 FISH 检测到的基因表达的层状分布。横线表示均值,error bars 表示 s.d 值,点表示单个数据点。n= 2 slices for BARseq2 and n= 3 slices for FISH。
f.使用 BARseq2 和 Allen 基因表达图谱中观察到的相对基因表达。每个点代表一个基因在层流深度为 100-µm 的 bin 中的表达。灰点表示层流位置随机数据之间的相关性。线性拟合和对角线和阴影区域显示 95% 置信区间。n= 2 slices for BARseq2 and n= 1 slice for ABA ISH。
g.听觉皮层 BARseq2 和 scRNA-seq 中每个细胞总读数的分布。只包括由 BARseq2 检测到的面板中使用的基因。
h.使用 BARseq2 或 scRNA-seq 检测到的每个基因的平均表达。每个点都代表一个基因。虚线表示 BARseq2 和 scRNA-seq 之间相同的表达。
i.在 BARseq2 和 scRNA-seq(紫点),或在两个 scRNA-seq 数据集(蓝点)中观察到的基因对之间的相关性。
j.Slc17a7 和 Gad1 在单个神经元中的表达。神经元主要表达 Slc17a7(蓝色)或 Gad1(红色),或两者均表达强烈(灰色)。
k.在两个 scRNA-seq 数据集中的神经元中,Slc17a7 和 Gad1 的排斥性指数(Methods),听觉或运动皮层中的 BARseq2 数据。
与之前的报告一致,许多钙粘蛋白在皮层的特定层和亚层中都很丰富 (Fig. 2d)。有趣的是,尽管大多数钙粘蛋白在听觉皮层和运动皮层中都有相似的层流表达,但一些钙粘蛋白在这两个区域的表达方式不同。例如,Cdh9 和 Cdh13 在听觉皮层中在 L2/3 富集,但在运动皮层中却不富集 (Fig. 2d and Extended Data Fig. 2)。钙粘蛋白峰值表达点的层流位置与其他方法一致,包括 RNAscope (Fig. 2e) 和 ISH 的 Allen Brain Atlas (ABA) 数据库 (Fig. 2f, Extended Data Fig. 3 and Methods)。因此,BARseq2 准确地解决了钙粘蛋白的层流表达模式。
然后,作者表征了 BARseq2 在单细胞分辨率下获得的基因表达 (Methods)。作者将听觉皮层中的 228,371 个 rolonies 分为 3,377 个兴奋性或抑制性神经元 (67.6±28.8(mean±s.d.) rolonies per neuron)在,运动皮层中的 752,687 个 rolonies 分为 11,492 个兴奋性或抑制性神经元 (65.5±26.0 (mean±s.d.) rolonies per neuron)。大多数钙粘蛋白在这两个皮质区域的单细胞表达水平上略有差异 (Extended Data Fig. 4)。在听觉皮层中,BARseq2 中每个细胞的总读数高于使用 10x Genomics v3 的单细胞 RNA 测序 (scRNA-seq) (Fig. 2g; median read count was 64 for BARseq2 (n=3,337 cells) compared to 57 for scRNA-seq (n=640 cells); P=5.3×10-5, rank-sum test)。因此,即使使用了有限数量的探针,BARseq2 也实现了至少等于 10x v3 scRNA-seq 的灵敏度。对于需要更好地量化低表达基因的实验,使用更多的探针可以进一步提高灵敏度。
进一步的分析表明,通过 BARseq2 检测 mRNA 具有特异性。用 BARseq2 测定的基因平均表达与用 10x v3 scRNA-seq 测定的基因平均表达高度相关 (Fig. 2h; Pearson correlation r=0.88)。一些异常值在 BARseq2 中的计数远比在 scRNA-seq 中要多,可能反映了细胞类型的采样差异、区域特异性基因表达和原位 RNA 可访问性的差异。例如,由 BARseq2 观察到的 Cdh6 的表达是由 scRNA-seq 观察到的 26 倍。这种差异可以归因于 scRNA-seq 数据中 Cdh6 表达的锥体束(PT)神经元的采样不足,以及 Cdh6 不同表达的邻近皮层区域的可能差异采样。此外,由 BARseq2 确定的单个神经元中基因对之间的相关性与使用 10x v3 scRNA-seq 一致,与两个独立的 10x v3 的实验相似 (Fig. 2i–k, Extended Data Fig. 5a,b and Methods)。这些结果表明,由 BARseq2 观察到的单细胞基因表达模式与 scRNA-seq 相当。
作者测试了 BARseq2 是否可以同时检测到更多的基因,从而能将投射与更大的基因集联系起来。因为 BARseq2 成像时间与检测到的基因数量成对数比例 (Fig. 1d),BARseq2 的多重容量不受成像时间的限制。此外,靶向多达 65 个基因并不显著影响每个基因的检测灵敏度 (Extended Data Fig. 5c and Methods)。在运动皮层中检测到这个 65 个基因集 (Fig.3a) 允许我们将神经元分类为由 scRNA-seq 定义的 9 个转录组神经元类型之一 (Fig. 3b, Methods and Extended Data Fig. 5d–h)。与之前的研究一致,这些转录组神经元类型显示出不同的层流分布 (Fig. 3b,c and Methods) 和钙粘蛋白的表达 (Fig. 3d)。大多数转录组类型都在预期的层中发现,除了 L5 PT 和 L6 IT Car3,它们出现在附加层(例如,L2/3)中。细胞分型中的这些不准确可能是由于标记基因的选择不理想造成的(see Methods for a detailed discussion),未来可能会通过优化基因集得到改进。然而,这些优化却超出了本研究的范围。这些结果表明,BARseq2 可以应用于探测由高于数十个基因组成的基因集,灵敏度降低最小,成像时间增加最小。
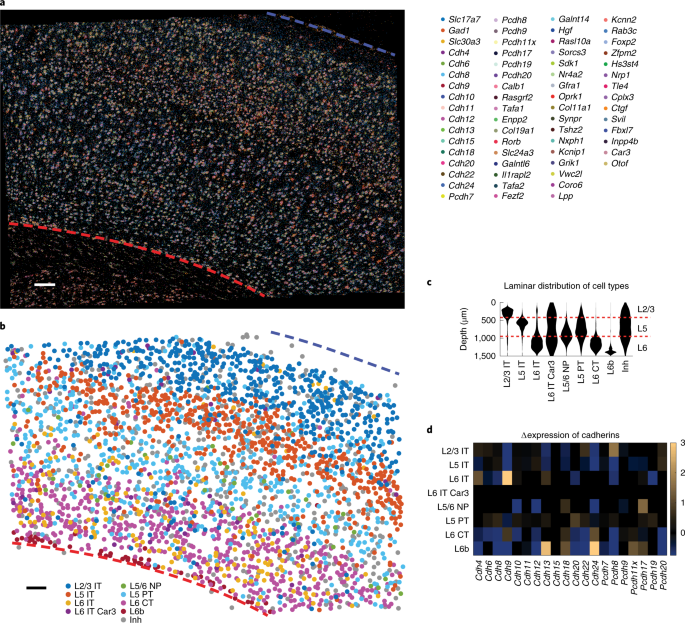
a.运动皮层中 rolonies 的代表性图像(来自四个切片测序)。mRNA 身份的颜色编码如图所示。大脑皮层的顶部和底部分别由蓝色和红色的虚线表示。比例条,100µm。
b.a 中显示的基于基因表达 call 出来的转录组细胞类型。
c.基于 BARseq2 观察到的标记基因表达的转录组神经元类型的层流分布。层标识显示在右边。NP,near-projecting。
d.由 BARseq2 鉴定的转录组神经元类型中钙粘蛋白的差异表达。过表达表示为黄色,表达不足表示为蓝色。仅显示了具有统计学意义的差异表达。通过对指示转录组类型和所有其他神经元类型中该基因表达之间的 two-tailed rank-sum test 来确定统计学意义。
3. BARseq2 将基因表达与投射相关联
先前对投射模式和基因表达之间关系的研究主要集中在揭示转录组神经元类型的投射模式上。虽然这种方法已经确定了某些转录组类型的投射模式,但 IT 神经元投射的多样性在很大程度上无法通过转录组类型来解释。为了进一步理解基因表达和投射之间的关系,作者提出了一种可选择的方法,成像一组与不同投射相关的靶向基因。这种方法依赖于 BARseq2 同时询问多个基因的表达和多个目标投射的能力,因此使用现有的转录组学方法很难实现,这些方法只能询问一个或少量的投射(例如,Retro-seq)或基于条形码的投射绘图方法,只能询问少数基因(例如,BARseq)。
作为一项原理证明研究,作者研究了远程轴突投射和三只小鼠运动皮层和听觉皮层中 20 种钙粘蛋白以及三个标记基因的表达。作者优化了 BARseq2,在不影响灵敏度的情况下在 barcoded 神经元内同时检测内源 mRNAs 和 barcodes (Extended Data Fig. 6a and Methods)。在每个条码细胞中,作者使用条形码测序图像来分割条形码胞体 (Fig. 4a)。然后将与这些像素重叠的内源性基因 rolonies 分配到 barcode 细胞 (Fig. 4a)。这使得我们能够在相同的神经元中同时绘制投射模式和基因表达 (Fig. 4b)。作者将这些目标位点的条形码与 15 个听觉皮层切片和 16 个运动皮层切片的 3164 个分割好的条形码神经元(1283个来自听觉皮层,1881个来自运动皮层)进行匹配,每一个厚度为10-µm。在条形码神经元中,624 个和 791 个神经元分别在听觉皮层和运动皮层的噪声层上方有投射。大多数听觉神经元(53%(329/624))和运动皮层神经元(89%(703/791))投射到大脑的多个区域。然后,作者重点研究了听觉皮层的 598 个神经元和运动皮层的 751 个神经元,这些神经元在每个细胞中也检测到足够的内源性 mRNAs,用于进一步分析 (Fig. 4c)。这些观察结果与之前在听觉和运动皮层进行的 BARseq 实验很大程度上一致,证实了对 BARseq2 的修改并没有影响投射绘制。
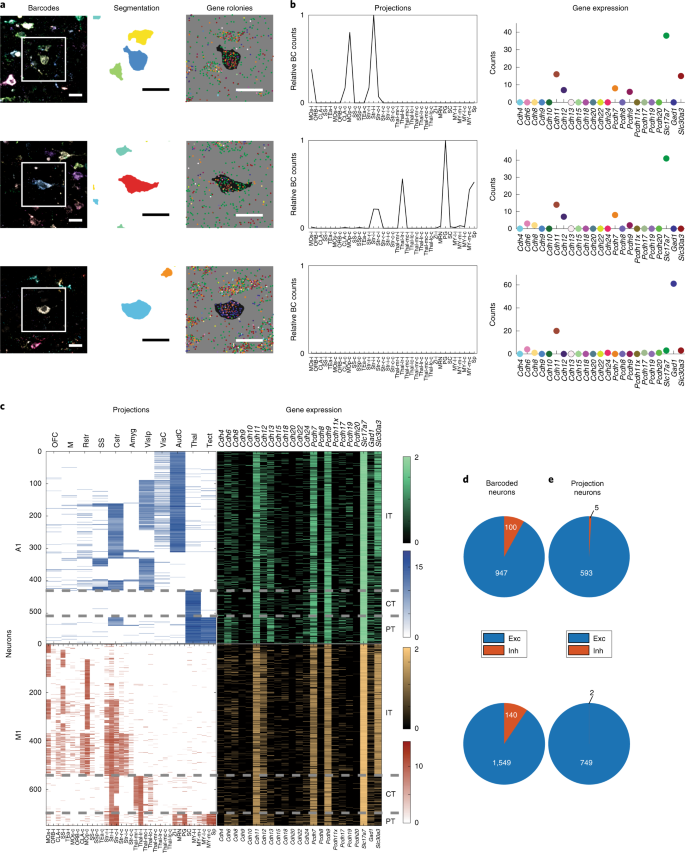
a.来自运动皮层的三种代表性神经元的伪彩色条形码测序图像、神经元胞体分割、基因 rolonies。分割和基因rolony 图像对应于条形码图像中的白色方框。在 gene rolony 图像中,与目标神经元的胞体分割相对应的区域是黑色的。比例尺,20µm。
b.a 中显示的目标神经元的投射和基因表达。表示基因表达的点使用与 a 中相同的颜色编码。前两排显示的神经元是兴奋性投射神经元,而下排显示的神经元是一个没有投射的抑制性神经元。每个小的目标区域对应的大脑区域见 Supplementary Table 2。BC, barcode。
c.听觉皮层(A1)和运动皮层(M1)中神经元的投射和基因表达。每一行都表示一个带条形码的投射神经元。投射和基因表达都用 log 显示。由投射模式确定的主要投射神经元类别在右边表示。
d-e.所有条形码神经元 (d) 或条形码投射神经元 (e) 中的兴奋性 (exc) 或抑制性 (inh) 神经元的数目。听觉皮层的神经元在上一行,运动皮层的神经元在下一行。
4. BARseq2 概括了已知的投射偏差
虽然 BARseq2 可以读出同一神经元中的基因表达和投射,但人们可能会担心,barcoding 神经元使用 Sindbis 病毒可能会破坏基因表达。为了确定基因和投射之间的关系,我们需要 Sindbis 感染的单个神经元的基因-基因关系与非感染的神经元进行对照,而绝对基因表达水平的任何变化都没有什么影响。值得肯定的是,先前的报道表明,尽管 Sindbis 感染的细胞中基因的绝对表达有所减少,但单个神经元中基因之间的关系确实在很大程度上保留了下来。此外,Sindbis-感染的神经元中转录组类型和投射之间的相关性被其他不需要 Sindbis 感染的方法所证实。与之前的报告一致,作者观察到,尽管基因表达整体减少,但条形编码神经元的相关性与非条形编码神经元的基因相关性一致(Extended Data Fig. 6b–f and Methods)。因此,基因表达与 BARseq2 解决的投射之间的关系可能反映了非条形码神经元中的这一点。
为了进一步测试 BARseq2 是否能捕获基因表达和投射之间的关系,作者询问是否可以识别转录组神经元类型的投射模式的差异,这也可以被以前的研究或其他实验技术验证。作者在三个不同的粒度级别上进行了这些验证分析。首先,BARseq2 证实了大多数具有长距离投射的条形编码神经元是兴奋性的,而不是抑制性的;有条形编码神经元的 8-9% 是抑制性的 (听觉皮层有 100/1047 个,运动皮层有 140/1689 个 Fig.4d),只有 7/240 个(3%) 的抑制性神经元中可以检测到的投射 (5 个在听觉皮层,2 个在运动皮层) (Fig. 4e, Methods and Extended Data Fig. 6g,h)。其次,BARseq2 鉴定了许多钙粘蛋白(8 个是听觉皮层,12 个是运动皮层),它们在 IT、PT 和丘脑皮质 (CT) 神经元中表达不同 (Fig. 5a–d);这些基因的差异表达与 scRNA-seq 的观测一致 (Extended Data Fig. 7a and Methods)。最后,BARseq2 证实了在听觉皮层中转录定义的 IT 亚型的投射模式中的已知偏差(Extended Data Fig. 7b,c and Methods)。因此,BARseq2 概括了 IT 神经元转录亚型之间已知投射差异。
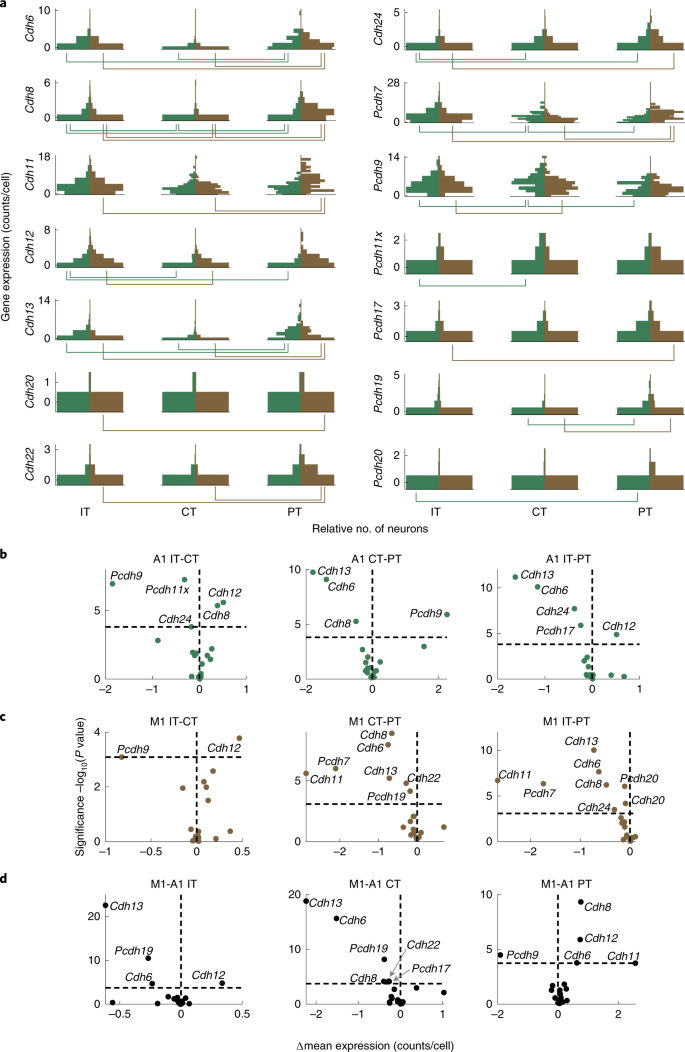
a.钙粘蛋白表达的垂直直方图(每个细胞的 raw counts),在听觉或运动皮层的主要类别中差异表达。y 轴表示基因表达水平(每个细胞的 counts),x 轴表示在该表达水平上的神经元数量。神经元的数量在图中被归一化,有最多神经元数目的 bins 具有相同的 bar lengths。在每个图中,听觉皮层(绿色) 的基因表达在左边,运动皮层(棕色)的基因表达在右边。每个图下的行表示不同基因表达的主要类对(FDR<0.05)。
b-c.钙粘蛋白在听觉皮层 (b) 或运动皮层 (c) 的成对主要类别中不同表达的火山图。y 轴表示显著性,x 轴表示效果大小。水平虚线表示 FDR<0.05 的显著性水平,垂直虚线表示相等的表达。
d.在指定的主要类别中,在听觉和运动皮层中差异表达的钙粘蛋白的火山图。y 轴表示显著性,x 轴表示效果大小。为了清晰起见,表示基因身份的近距离的点会被标注为灰色箭头。水平虚线表示 FDR<0.05 的显著性水平,垂直虚线表示相等的表达。对于所有 panels,使用 two-tailed rank-sum tests 计算 P 值。
5. BARseq2 确定了钙粘蛋白与 IT 投射的相关性
在确定了 BARseq2 识别了与先前研究一致的投射的基因相关性后,作者询问钙粘蛋白的表达是否与 IT 类神经元中的投射模式有相关性。虽然钙粘蛋白和其他细胞粘附分子在发育期间参与了投射规范和轴突生长,但许多分子在后期发育阶段发挥了与投射规范无关的其他功能。此外,其他机制,如轴突剪枝,可以进一步塑造神经元的投射模式,独立于最初的基因程序。因此,钙粘蛋白和投射之间的任何相关性都可能是最初发育程序的残余,或“再现”,因此可能很弱,并进一步被与后期发育阶段相关的基因表达所掩盖。为了克服识别基因表达和投射之间潜在的弱关系的挑战,作者使用 BARseq2 来识别投射和钙粘蛋白之间的相关性,使用一种基于模块的策略,受转录组学中类似方法的启发。在测量单个投射和基因时,投射模块和基因模块平均超过噪声,因此在测量中有相当大的生物和/或技术噪声时更容易检测到。这种方法需要了解单个神经元对许多大脑区域的投射,这是与基于条形码的投射绘图技术相比,基于条形码的投射绘图技术 (即 BARseq 和 BARseq2) 的一个独特优势。接下来,作者确定了钙粘蛋白的表达和 IT 神经元中的投射之间的适度关联,包括跨皮层区域共享的一些相关的钙粘蛋白/投射对。
IT 神经元对其靶点的投射不是随机的。相反,在听觉皮层和运动皮层中,这些投射都是有组织的,并显示出统计规律,可以在 BARseq 获得的大数据集中被发现 (Fig. 6a)。例如,听觉皮层中投射到躯体感觉皮层 (somatosensory cortex,SS) 的神经元也更有可能投射到同侧视觉皮层 (ipsilateral visual cortex,Vislp),而不是投射到对侧听觉皮层 (contralateral auditory cortex,VisC)。为了利用这些相关性,作者使用非负矩阵分解 (NMF) 将每个神经元的投射模式表示为几个“投射模块”的总和。(NMF 是一种与主成分分析相关的算法,但施加了附加的非负投射约束)。这些模块中的每一个(运动皮层的六个模块和听觉皮层的三个模块;Fig. 6b)由可能同时出现的投射子集组成。作者通过主要投射(皮层(CTX)或纹状体(STR))和投射的侧面(同侧(I)或对侧(C))命名这些模块。对于一些模块,作者进一步指出,通过前缀“C”(例如,CSTR-I 或 CCTX-I),投射是在结构的尾部部分。少量的投射模块可以解释投射中的大部分差异 (3个模块和6个模块分别解释了听觉皮层的9个区域和运动皮层的18个区域的84%和87%的投射差异;Fig. 6c)。
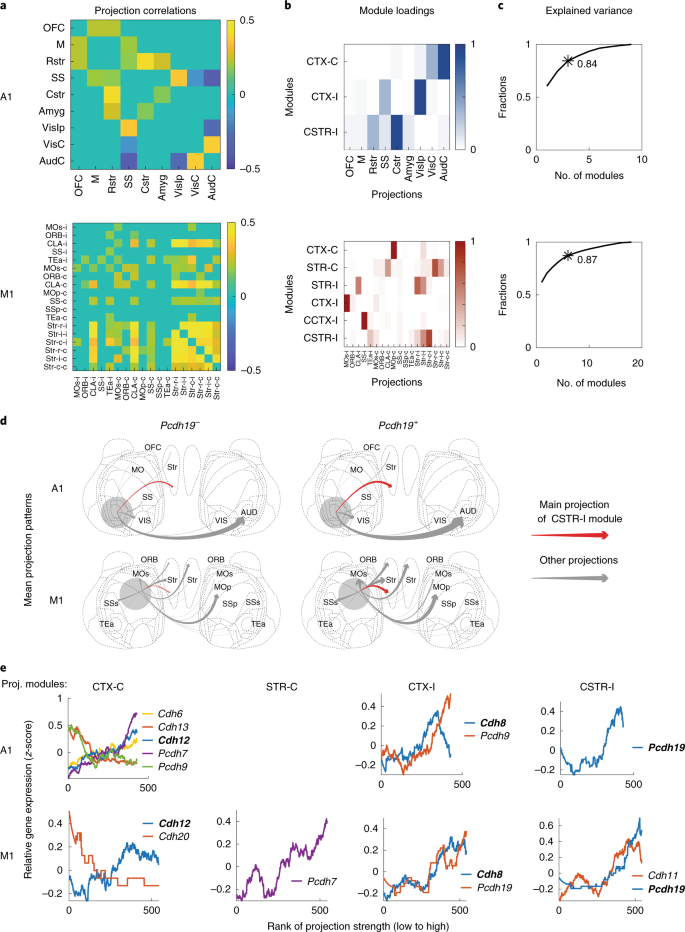
a.听觉皮层(A1)或运动皮层(M1)IT 神经元中大脑不同区域投射的皮尔逊相关性。只显示出显著的相关性。OFC:眶额叶皮层、M:运动皮层;Rstr:股侧纹状体、SS:躯体感觉皮层;Cstr:尾部纹状体、Amyg:杏仁核、VisIp:同侧视觉皮层;ViSC:对侧视觉皮层;AudC:对侧听觉皮层。
b.听觉皮层(顶部)或运动皮层(底部)中 IT 神经元的投射模块。每一行都代表一个投射模块。列表示对不同大脑区域的投射。
c.听觉皮层(顶部)和运动皮层(底部)不同投射模块解释的方差分数。与 b 中对应的投射模块的数量用星号标记,带有方差分数说明。
d.A1 和 M1 中神经元的有无 Pcdh19 表达的平均投射模式。箭头的厚度表示投射强度(条形码计数)。红色箭头表示对应于 CSTR-I 投射模块中最强投射的投射。ORB:眶额叶皮层;MO:次级运动皮层;MOp:初级运动皮层;SSp:初级躯体感觉皮层;SSs:二级躯体感觉皮层;TEa:时间关联皮层。
e.钙粘蛋白(y轴)的表达与听觉皮层和运动皮层中指示的投射模块的排名相关。神经元(x轴)根据指示的投射模块的强度进行排序。仅显示了与投射模块显著相关的基因 (使用 FDR<0.1 的 two-tailed rank-sum tests)。这两个区域与相同投射模块相关的基因用粗体显示。
由于神经元的投射模式及其转录组类型都与层状位置有很好的相关性,作者研究了,与神经元的层状位置相比,钙粘蛋白如何解释 IT 神经元投射的多样性 (Methods)。虽然大多数钙粘蛋白在投射模块上没有预测能力,但与神经元的层流位置所解释的差异相比,一些钙粘蛋白可以解释投射差异的很大一部分 (Extended Data Fig. 8)。比如 Cdh13 和 Pcdh7 解释了 6.0%±0.3% 和 7.0%±0.3%(平均值±标准差)听觉皮层中 CTX-C 的变化,相比之下,神经元的层状位置解释了 19.4±0.3%(平均值±标准差)。引人注目的是,Pcdh19 和 Pcdh7 可以预测听觉皮层中的 CSTR-I,而层流的位置则不是。这些结果表明,一些但不是所有的钙粘蛋白都是对投射的适度预测,而这些钙粘蛋白的预测能力在数量上可以与神经元的层流位置相比较,已知的最强的投射模式预测因子之一。
为了进一步理解钙粘蛋白表达如何与投射的关系,作者研究了它如何与投射模块共变化(Supplementary Fig. 1)。有趣的是,几种钙粘蛋白的表达在两个皮质区域与相似的投射模块共变化。例如,听觉皮层中表达 Pcdh19 的神经元比不表达 Pcdh19 的神经元投射到 CSTR-I 更强(Fig. 6d;P=5×10−4 使用 rank-sum test 比较神经元中有(n=83)或无(n=346)Pcdh19 表达的 CSTR-I 模块)。同样,Cdh8 与 CTX-I 模块相关,Cdh12 与 CSTR-I投射模块之间的关联也在运动皮层中看到(Fig. 6d;P = 4 × 10−6 using rank-sum test, n = 31 for Pcdh19+ neurons and n = 512 for Pcdh19− neurons)。同样地,Cdh8 与 CTX-I 模块相关,Cdh12 与 CTX-C 模块相关 (Fig . 6e;false discovery rate (FDR) < 0.1)在听觉和运动皮层。通过使用 cholera toxin subunit B (CTB) 和 FISH 的逆行追踪,独立地验证了这些相关性 (Extended Data Fig. 9a–e and Methods)。Pcdh19 和 Cdh8、Cdh11,分别与运动皮层中的 CTX-I 或 CSTR-I 模块相关 (Fig. 6e and Extended Data Fig. 8),与钙粘蛋白的潜在组合性质的预测一致。虽然单个钙粘蛋白和投射之间的相关性相对适中,但作者观察到,相同的钙粘蛋白在这两个领域与相似的投射模块相关,表明共同的分子逻辑可能强调皮层区域投射的组织。
基于单个基因表达的分析存在单个神经元中基因表达的生物学和技术噪声。作者推断,基因之间的相关性可能允许我们通过分别分析每个基因来识别基因表达和投射之间的额外关系。这种利用基因之间关系的能力代表了 BARseq2 比原始 BARseq 的优势,因为 BARseq2 提高了多重基因检测的能力。为了利用基因之间的相关性,作者根据运动皮层中 IT 神经元的 7 个 scRNA-seq 数据集,将 16 个钙粘蛋白分为 3 个元分析共表达模块 (Fig. 7a and Extended Data Fig. 10a,b)。为了获得模块,作者遵循 Ballouz 等人和 Crow 等人定义的基于等级的网络聚合程序,将七个特定于数据集的基因-基因共表达网络组合成一个聚合网络,然后使用动态树切割算法将显示一致过度相关的基因组合在一起。两个共表达的模块与投射相关:模块 1 与对侧纹状体投射相关(STR-C 投射模块),而模块 2 与同侧尾纹状体投射相关 (CSTR-I; Fig. 7b,c and Extended Data Fig. 10c,d)。共表达模块和投射之间的这些关联与每个模块中包含的单个基因和相同投射之间的关联一致,但比强度更强 (Extended Data Fig. 10e)。有趣的是,这些共表达模块在 IT 神经元的多个转录组亚型中都很丰富,但这些转录组亚型在转录组分类学的多个分支中被发现 (Fig. 7d and Extended Data Fig. 10f)。例如,模块 1 与L2/3、L5 和 L6 中的 IT 神经元的转录组亚型相关联。这一结果与之前的观察结果一致,即 IT 神经元的第一层转录组亚型(即 IT 类中转录组分类中最高级别的亚型)似乎共享投射模式,并进一步提高了转录组分类不必须捕捉投射的差异。总之,作者发现投射与独立于转录组亚型的钙粘蛋白共表达模块相关,这表明 BARseq2 可以揭示基因表达和投射模式之间的复杂关系。
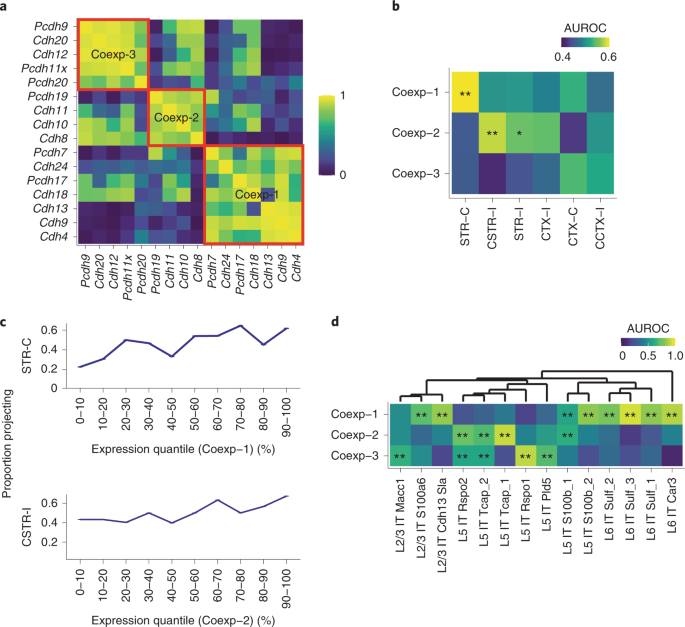
a.在运动皮层的 IT 神经元中使用 scRNA-seq 识别的钙粘蛋白之间的相关性。三个共表达式模块由红色方框标记。不显示不属于任何模块的钙粘蛋白。
b.钙粘蛋白共表达式模块与投射模块之间的关联(AUROC, area under the receiver-operator characteristic curve)。显著的关联由星号标记(*FDR<0.1,**FDR<0.05)。
c.具有指示投射模块的神经元部分作为共表达模块表达的函数。神经元被基因模块分位数 binned。
d.single-cell SmartSeq 数据集中转录组 IT 神经元中三个共表达模块的关联(AUROC,显著性如 b 所述)
讨论
BARseq2 结合了对许多大脑区域的投射的高通量绘图和在单细胞分辨率下的基因表达的多重检测。由于 BARseq2 是高通量的,我们能够在一个实验中关联数千个单独神经元的基因表达和投射模式,从而实现使用其他单细胞技术具有挑战性的统计能力。通过将 BARseq2 应用于成年小鼠的两个遥远的皮层区域——初级运动和听觉皮层,确定了不同投射的钙黏蛋白相关性。作者的结果表明,BARseq2 提供了一条途径来发现整个皮层共享的基因表达和投射的一般组织。
通过 BARseq2 进行高通量多重基因检测
为了将基因组与投射相关联,作者设计了 BARseq2 来以高通量检测基因表达,用于对数十个基因进行多重检测,以具有足够的灵敏度,并与基于条形码的投射映射兼容。为了满足这些需求,作者将 BARseq2 基于锁式探针的方法。通过对灵敏度、测序读数以及与条形码测序的兼容性进行额外优化,成功地使用 BARseq2 来识别投射的基因相关性。
BARseq2 的关键要求之一是读取许多基因时的高通量。通过使用 Illumina 测序化学技术对 mRNA 进行强力扩增、组合编码和强大的读出,与许多其他基于成像的空间转录组学方法相比,BARseq2 可以在低光学分辨率下实现快速成像。进一步的优化,包括解决空间混合 rolonies 的计算方法,有可能进一步提高成像通量。尽管 BARseq2 的基因复用能力最终可能会受到其他物理限制的限制,例如 rolonies 的拥挤和检测灵敏度降低,但当复用数十到数百个基因时,这些因素不太可能受到限制。
另一个关键的优化是提高早期版本的基于锁式探针的技术容易受到的低灵敏度影响,除非使用特殊且昂贵的引物。受其他空间转录组学方法的启发,作者和其他人发现用多个探针平铺靶基因可以大大提高灵敏度。该设计允许针对不同的实验目的改变灵敏度。尽管在目前的工作中,作者仅使用每个基因适量的探针来识别钙粘蛋白与投射的相关性,以达到与使用 10x Genomics v3 的 scRNA-seq 相似的灵敏度,但当使用更多探针时,BARseq2 的灵敏度可能会高得多(Fig. 1e)。这种高且可调的灵敏度,再加上 BARseq2 的基因复用能力不受成像时间限制,为 BARseq2 的潜在应用开辟了解决需要高通量原位基因表达询问的广泛问题的机会。
BARseq2 揭示了预测的基因相关性
BARseq2 利用 BARseq 提供的高通量轴突投射映射来识别不同投射的基因相关性。BARseq 的灵敏度与单神经元追踪相当。尽管 BARseq 投射的空间分辨率低于传统的单神经元追踪技术,但它提供的通量比最先进的单细胞追踪技术高几个数量级。这种高通量使 BARseq 能够揭示投射模式中的高阶统计结构,而使用现有技术(例如单细胞追踪)很难观察到这些结构。BARseq 统计能力的提高是以一定空间分辨率为代价获得的,让人想起不同通量和读取深度的 scRNA-seq 技术的不同聚类能力。因此,BARseq 的高通量为研究投射模式的组织及其与基因表达的关系提供了强大的资产。
BARseq2 能够以单神经元分辨率和其他方法难以实现的规模同时测量多个大脑区域的多重基因表达和轴突投射。例如,Cre 依赖性标记允许询问基因定义的神经元亚群的基因表达和投射模式。然而,这种方法缺乏细胞分辨率,受到 Cre 细胞系可用性的限制,并且需要通过一个或两个基因的表达来具体区分感兴趣的神经元群体。单细胞转录组技术与逆行标记的结合确实提供了细胞分辨率,但一次只能询问对一个或最多少数大脑区域的预测。无法询问同一神经元对许多大脑区域的投射会错过投射中的高阶统计结构,这些结构是非随机的,并提供有关神经元其他属性的附加信息,例如层状位置和基因表达。可以使用多重单细胞追踪来获得单个神经元到多个大脑区域的投射,但这些方法的通量仍然相对较低。此外,许多先进的单细胞追踪技术需要特殊的样品处理,这阻碍了对同一样品中基因表达的多重询问。原始 BARseq 解决了单细胞投射绘制的通量问题,但可与投射共同询问的基因数量较少(最多三个)限制了其在识别基因表达与投射之间的一般关系中的使用。因此,BARseq2 解决了现有技术的局限性,并提供了一种强大的方法来探测基因表达和投射模式之间的关系。
钙粘蛋白与 IT 神经元的不同投射相关
作为一项原理验证研究,作者使用 BARseq2 来识别与听觉皮层和运动皮层中同源 IT 投射相关的几种钙粘蛋白,这两个空间和转录组上遥远的区域具有不同的皮层和皮层下投射目标。此外,与投射相关的钙粘蛋白共表达模块与转录组分类学的多个分支相关。神经元连接性与独立于转录组类型的基因表达变化之间的这种相关性并不是皮质所独有的,并且之前已在其他大脑区域(例如海马体)中观察到。因此,作者的研究结果与以下假设一致:共享的细胞粘附分子代码可能是独立于转录组类型的皮质投射多样性的基础。
尽管一些钙粘蛋白预测投射的能力在大小上与层流位置(投射模式的强有力预测因子)相当,但这些钙粘蛋白只能解释投射总体方差的一小部分。钙粘蛋白表达和投射模式之间的这种嘈杂关联与已知的钙粘蛋白在指定皮层和其他环路中的神经元连接中的作用形成对比,但由于某些原因,这些关联的相对较小的幅度并不令人惊讶。首先,指定投射所需的基因表达程序和信号线索在发育过程中通常是短暂的,因此这些钙粘蛋白很可能仅代表建立投射的常见发育程序的残余物,或者可能是持续功能或维持投射所需要的。其次,非钙粘蛋白细胞粘附分子(例如 IgCAMs)和其他细胞表面分子(例如 plexins、semaphorins 和 teneurins)也参与指定投射,因此钙粘蛋白可能只代表分子程序的一小部分指定投射。最后,皮质投射在初始规范后经历了广泛的活动依赖性修改,因此皮质投射的总体多样性可能远高于初始分子程序产生的多样性。通过应用 BARseq2 结合微扰实验来揭示投射神经元及其在发育过程中投射到的区域中的基因表达,可以更好地解决这些可能性。因此,BARseq2 提供了一条发现无数遗传程序的途径,这些遗传程序指定和/或与发育中和成熟动物的长程投射相关。
BARseq2 构建神经元多样性的统一描述
神经元条形码最初被提出作为一种在突触分辨率下解开环路连接的方法。通过条形码测序解决神经元连接问题,不仅有可能通过利用测序技术的进步来实现高通量和单细胞分辨率,而且还提供了一条在单个神经元中整合多个神经元特性测量的途径——迈向“Rosetta brain”。BARseq2 是朝着这一目标迈出的一步。虽然 BARseq2 目前只能以相对较低的空间分辨率(大脑区域,即数百微米)解析投射,但未来可以通过使用原位测序读取轴突条形码来解决这一限制(Yuan et al.,未发表的数据),这将以亚细胞空间分辨率解决轴突投射。进一步将轴突条形码的原位测序与突触标记、扩展显微镜和/或跨突触病毒标记相结合,可以产生有关神经元突触连接的信息。由于 BARseq2 使用空间信息整合神经元特性,因此它可能与其他原位测定兼容,例如免疫组织化学、双光子钙成像和树突形态重建。通过在空间上关联单个神经元中的各种神经元属性,BARseq2 代表了实现神经元环路全面描述的可行途径。
注:本文为个人学习笔记,仅供大家参考学习,不得用于任何商业目的。如有侵权,请联系作者删除。

本文由 mdnice 多平台发布