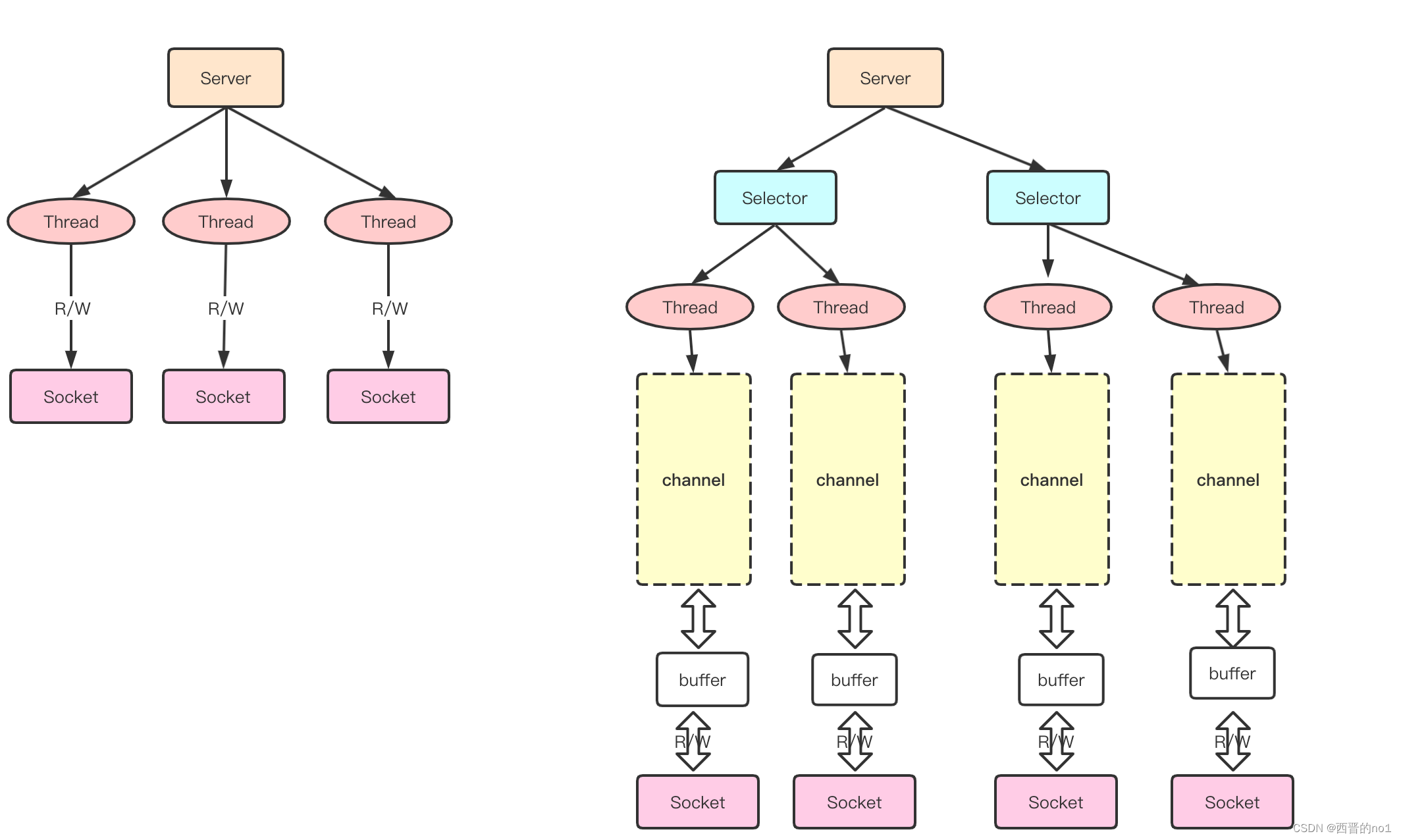
氧化应激是细胞有氧代谢的一个特征,在胃癌(GC)的发展和转移过程中起着至关重要的调控作用。长非编码 RNA(lncRNA)是胃癌发展过程中的重要调控因子。然而,目前对氧化应激相关lncRNAs(OSRLs)的预后模式及其在免疫微环境中的功能的研究尚不充分。
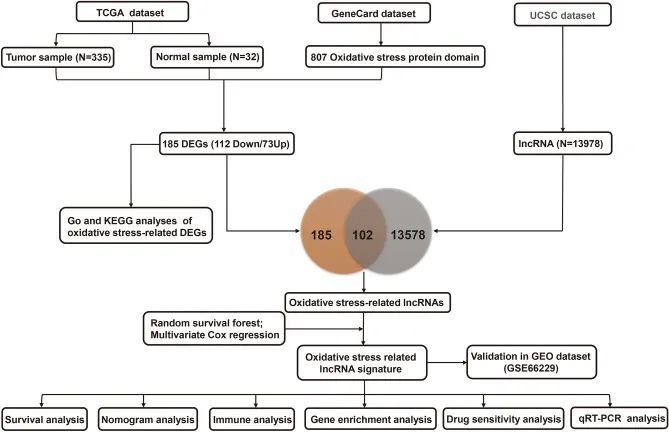
图1 本研究的流程图
1. 氧化应激相关基因的富集分析
在 TCGA-STAD 数据集中共发现了 185 个氧化应激相关 DEGs(73 个上调基因和 112 个下调基因)(图 2A )。然后,作者进行了 KEGG 分析(图 2B)和 GO 分析(图 2C ),以确定与氧化应激相关的 DEGs。KEGG分析的前5条通路分别是脂质与动脉粥样硬化、药物代谢-细胞色素P450、IL-17信号通路、化学致癌-受体激活、流体剪切应力和动脉粥样硬化。GO 分析包括生物过程(BP)、细胞成分(CC)和分子功能(MF)类别。在 BP 类别中,DEGs 富集于氧化应激反应、异生物刺激和 ROS 代谢过程。在CC类别中,DEGs与内囊腔、膜筏、膜微域等相关。在 MF 类中,DEGs 与受体配体活性、信号受体激活剂活性等有关。
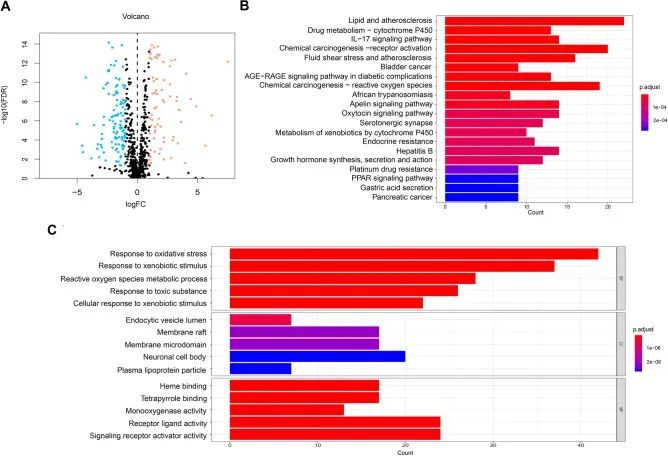
图2 氧化应激相关基因的差异表达基因鉴定和功能富集分析
2. OSRLs 预后特征的确定和验证
通过从 UCSC 数据集中提取 185 个氧化应激相关 DEGs 和 13,978 个 lncRNAs 的交集,作者发现了 102 个氧化应激相关 lncRNAs。随机生存森林算法显示,33 个 lncRNA 与氧化应激有相互作用。多变量分析证实 AC091057.1、TP53TG1、ARRDC1-AS1、SNHG5、DUXAP8、DIP2A-IT1、AL355001.1 和 COLCA1 为 OSRLs 预测特征。当风险评分高于中位值时,患者被归类为 HRiG;反之,则被归类为 LRiG。生存状态曲线(图 3A)和风险评分曲线(图 3C)显示亚组患者存在明显差异。KM 曲线显示,HRiG 患者的总生存率低于 LRiG 患者(P < 0.01)(图 3E)。此外,接收者操作特征(ROC)曲线和曲线下面积(AUC)值表明,风险模型在预测 GC 预后方面具有敏感性和特异性。在训练组中,3 年和 5 年的 AUC 值分别达到了 0.712 和 0.737(图 3G)。在结果相似的验证组中(图 3B、D、F),3 年和 5 年的 AUC 值分别达到 0.612 和 0.628(图 3H)。
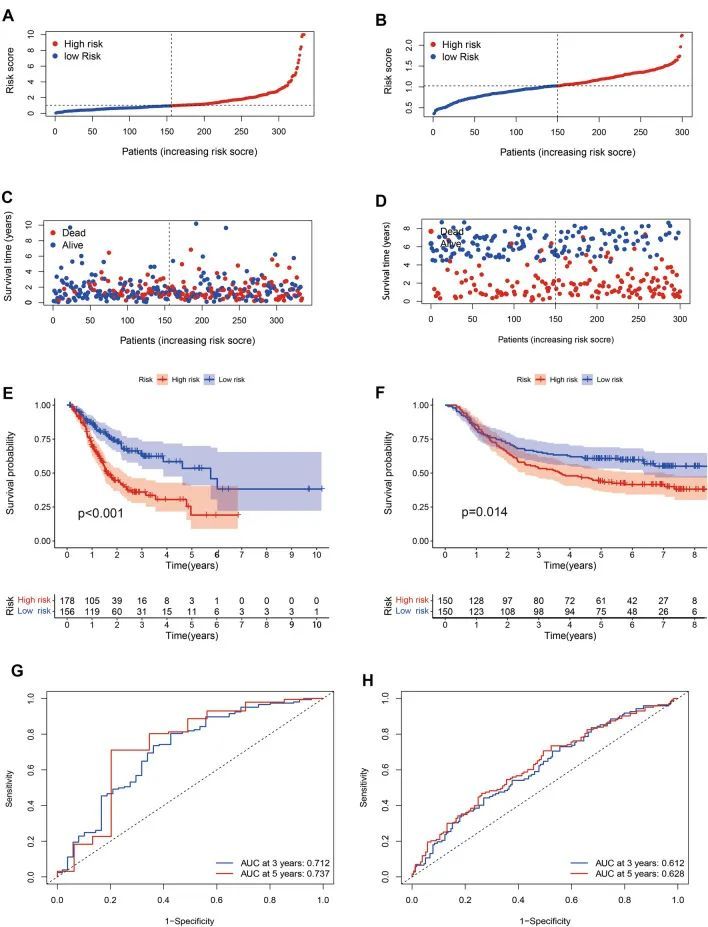
图3 OSRLs 特征的构建和验证
3. OSRL 特征的预后价值
值得注意的是,风险模型在两个单变量(HR = 1.156,95%CI 1.061-1.261)和多变量分析(HR = 1.133,95%置信区间1.033–1.243)(图4A、B),风险模型的AUC值达到0.737(图4C)。后来,作者利用OSRLs特征构建了一个预后列线图,以可视化每个患者的生存率,并评估了GC患者的3年和5年总生存概率(图4D)。使用校准曲线来评估列线图的预测性能。图4E和F显示,校准曲线的预测值和真实值呈线性相关。上述结果表明,预测特征和列线图在GC预测中具有良好的性能、准确的预测和判别能力,可在临床管理中发挥重要作用。
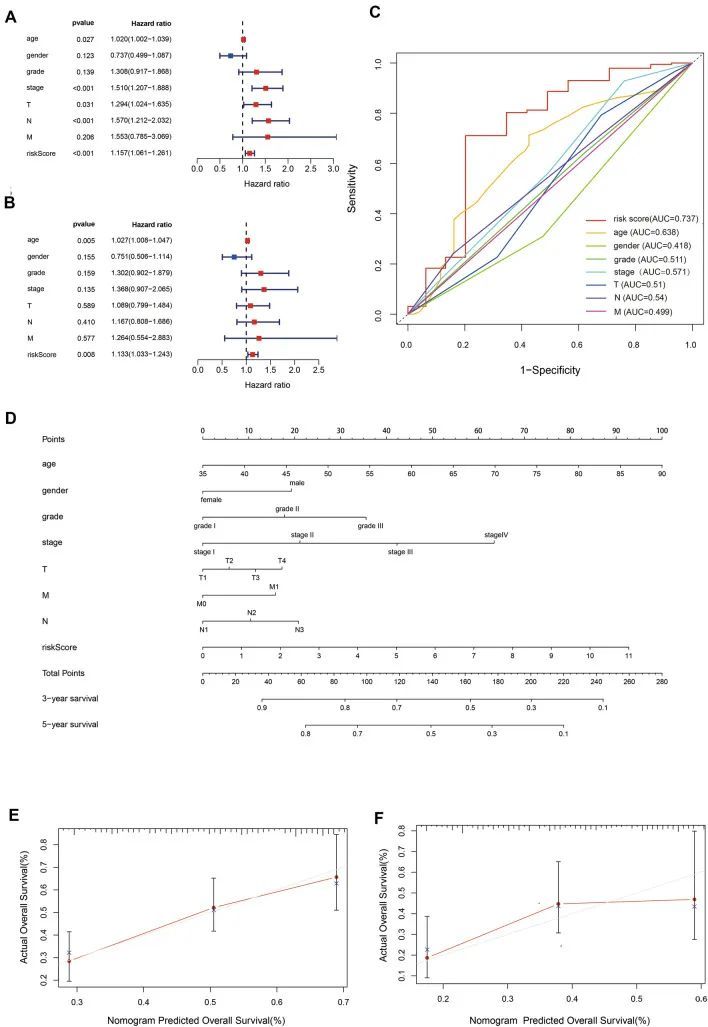
图4 OSRLs 特征的预后价值
4. OSRLs 特征的预后价值
预测特征的基因富集分析表明,LRiG 和 HRiG 患者主要富集于氧化应激和免疫通路。例如,缺氧标志基因(图 5A、D)代表在低氧水平下上调的基因,这些基因在 HRiG 中富集。活性氧通路(图 5B)在 LRiG 中富集;相关基因包括关键基因硫氧还蛋白(TXN)(图 5E),它与其他酶一起构成了参与调节线粒体 ROS 和保护细胞免受氧化应激的清除酶系统 18 。作为泛醌氧化还原酶亚基 A6(NDUFA6)的上游基因,TXN 可能会通过调节 NDUFA6 的表达来影响线粒体的活力 19 。标志性 Wnt β-catenin 信号传导(图 5C、F)和标志性 MTORC1 信号传导(图 5G、J)通路是 LRiG 中显著富集的免疫相关通路。此外,通过GSEA进行的KEGG通路分析显示,细胞因子-细胞因子受体相互作用(图5H,K)和细胞周期(图5I,L)分别在HRiG和LRiG患者中富集。
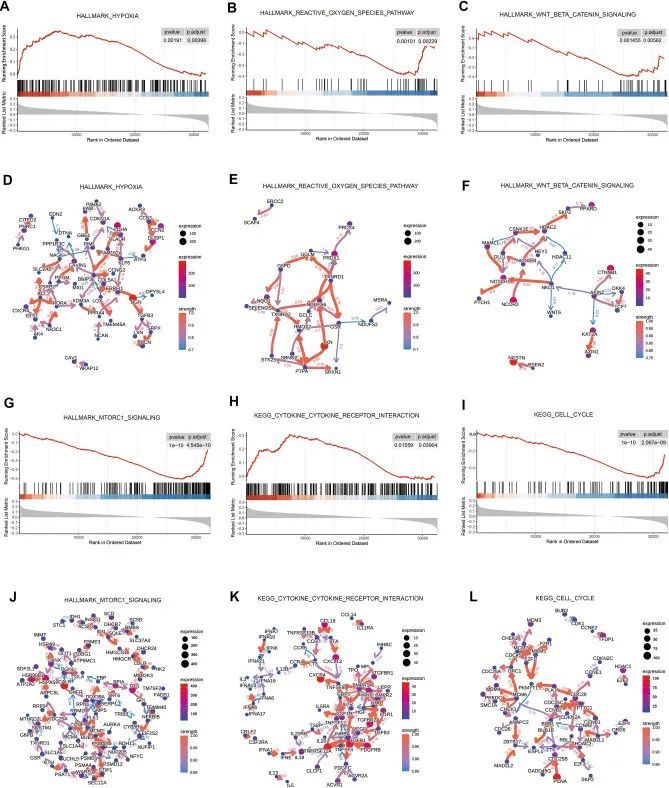
图5 通过 GSEA 对 OSRLs 特征进行通路富集分析
5. 肿瘤免疫微环境分析
为了证实OSRLs与肿瘤免疫微环境之间的相互作用,作者利用 "ssGSEA "软件包分析了亚组中16种免疫相关细胞和13种免疫功能评分的差异。如图 6A 所示,肥大细胞作为参与免疫反应的细胞,在不同亚组之间存在显著的富集差异(P<0.05)(图 6B)。主要组织相容性复合体 I 类(MHC I 类)在 LRiG 中的表达水平高于 HRiG(P<0.05)(图 6C)。这可能是因为 MHC I 类有助于 CD8 + T 细胞消灭恶性细胞并提供长期保护性免疫。与 MHC I 类的结果相反,副炎症(parainflammation)是组织细胞在各种压力或异常功能下的炎症反应,在高危患者中富集程度更高(P<0.05)(图 6D)。随后,利用热图估算了免疫检查点与风险模型之间的交互作用(图 6E),CD200、TNFSF4、TNFSF9 和 BTLN2 检查点的表达在 HRiG 患者中显著增加(图 6F-I)。最后,评估了探索OSRLs预测特征与化疗药物之间关系的测量标准(半数最大抑制浓度,IC 50 )。HRiG患者对甲氨蝶呤和TrKA抑制剂更敏感(图7A,B),而LRiG患者对芦卡帕利、bryostatin、embelin和palbociclib更敏感(图7C-F)。
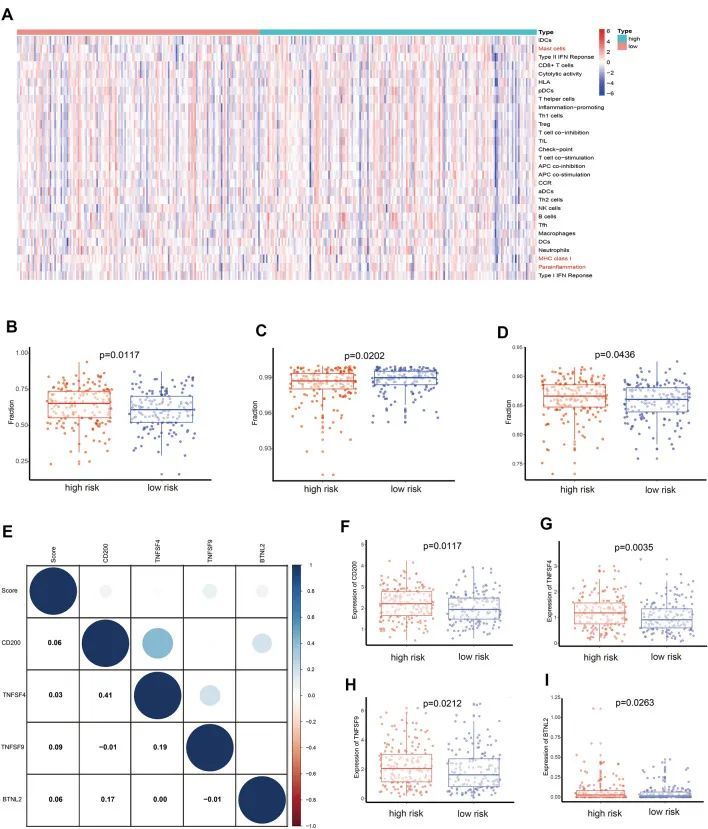
图6 根据 OSRLs 特征预测免疫细胞浸润
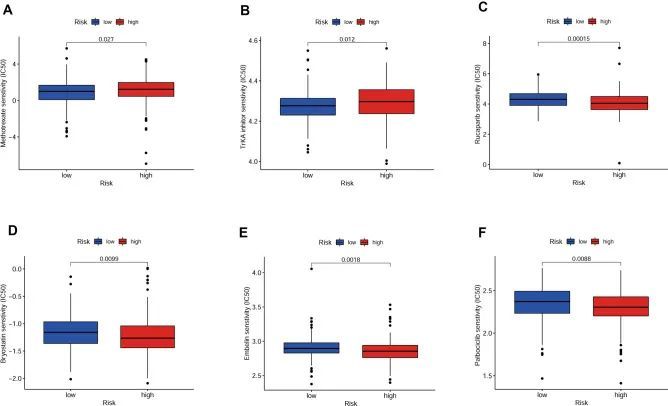
图7 基于OSRLs特征的化疗药物敏感性预测
6. 基于OSRLs特征的化疗药物敏感性预测
8个lncRNA的相对表达在AGS、HGC-27和GES-1细胞系中通过qRT-PCR实验得到了验证。如图 8A-G 所示,与 GES-1 细胞相比,DUXAP8、TP53TG1 和 AL355001.1 在 AGS 和 HGC-27 细胞中明显上调,而 AC091057.1 和 SNHG5 则明显下调。ARRDC1-AS1和COLCA1在HGC-27细胞中上调,而AGS细胞与GES-1细胞相比无明显差异。DIP2A-IT1的表达在GES-1、AGS和HGC-27细胞系中没有明显差异。
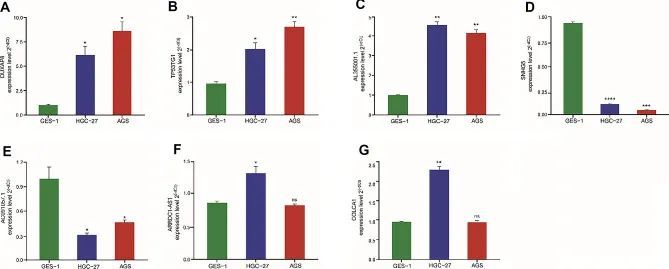
图8 7 种 lncRNA 在 GC 细胞系中的表达水平比较
总结
总之,经 qRT-PCR 实验验证的 OSRLs 有可能成为预测 STAD 患者总生存率的生物标志物。值得注意的是,该特征可能会调节免疫浸润水平和免疫功能。因此,氧化应激、lncRNAs、免疫和 GC 之间的机制和关系值得进一步探讨和验证。作者相信,8个OSRLs特征可以指导对GC生物学行为及其临床预后的研究。然而,本研究也存在一些局限性:(1)本研究使用了一个外部验证集,需要更多的外部验证集来确保模型的有效性。(2)有必要通过实验进一步验证 OSRLs 在 GC 中的作用机制。