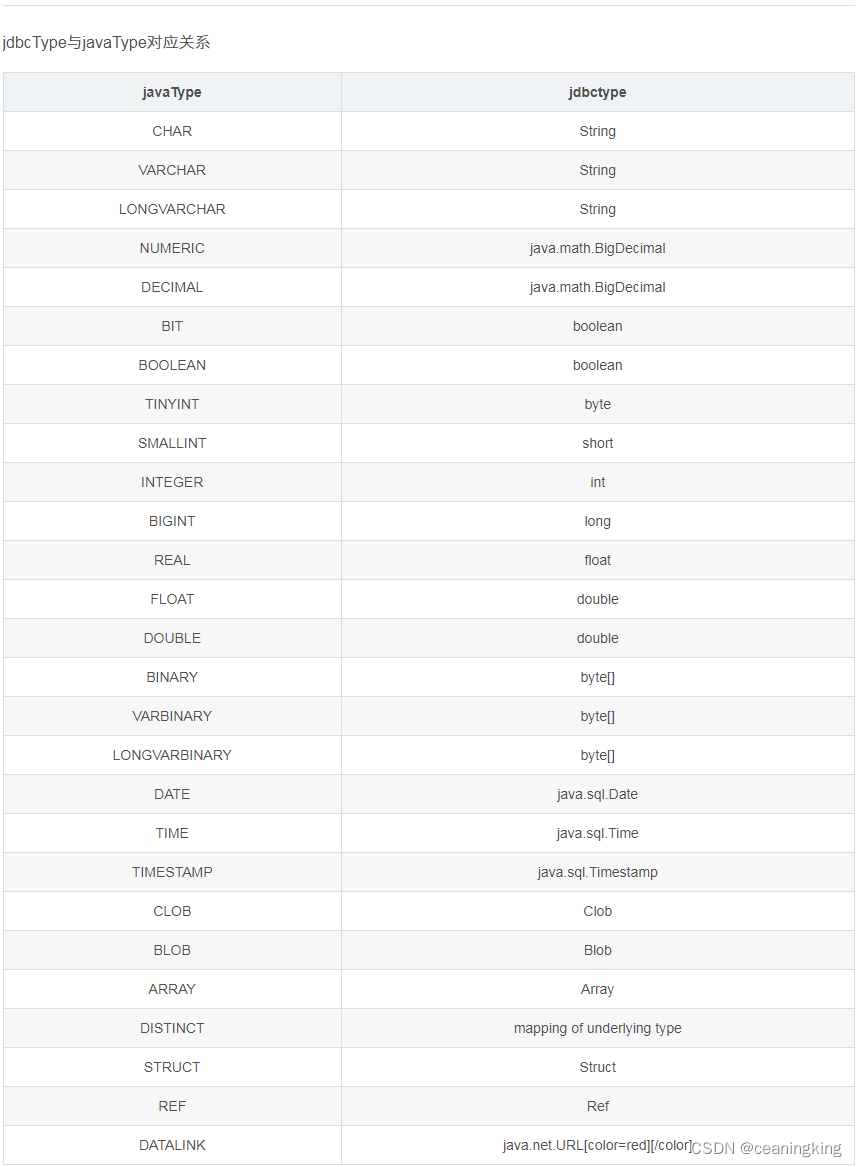
目录
近交与杂交的遗传学效应
(1) 近交使基因纯和,杂交使基因杂合
近交效应:
(2) 近交系数与亲缘系数
(3)运用通径分析方法计算近交系数和亲缘系数
① 通径与通径链
② 通径分析的理论及其应用
(4)近交降低群体基因型值的平均值,杂交提高群体均值
(5)近交使群体分化,杂交使群体一致
(6)近交与人工选择相结合使提高杂种优势的重要手段之一
杂种优势及其遗传理论
(1) 概念:
(2)杂种优势形成的机制有两种学说:
① 显性说(dominance hypothesis, Bruce, 1910)
② 超显性说(overdominance hypothesis)
近交与杂交的遗传学效应
非随机交配
- 理想群体假设群体中的交配是随机的,但很多群体对于某些性状而言交配并不是随机的。当非随机交配发生时基因型存在的比例就难以用哈迪一温伯格定律来进行估计
- 一类非随机交配是正选型交配(positive assortative mating)即带有相似表型的个体优先交配
- 在自然群体中这种情况较为普遍。例如人类的选择性就很高高个子女人和高个子男人婚配,矮个子女人与矮男人婚配都比较常见,比随机频率要高。
- 异型交配(nonassortative mating):基因型不同的纯合子之间的交配
- 同型交配 (assortative mating):相同基因型之间的交配
- 近交 (inbreeding):也称近亲繁殖或近亲婚配,是有亲缘关系的个体相互交配,繁殖后代。近亲繁殖按亲缘关系的近、远程度一般可分为:全同胞 (full-sb) (同父、母的兄妹)、半同胞 (half-sib) [同父(或母)(或)的兄妹]和表兄妹(first cousins)之间的交(婚)配
- 自交(selfing):植物的自花授粉、动物的自体受精(self-fertilization)自交是近亲繁殖中最极端的方式。
(1) 近交使基因纯和,杂交使基因杂合
以一对等位基因为例,有三种交配类型:

在AA × AA,aa × aa交配类型→产生的子代的基因型(或表型)全部都是与亲本相同的纯合体;
第三种交配类型AaxAa属于杂合体间的同型交配,其后代为1/4AA+1/2Aa +1/4aa,经过n代连续的同型交配的过程和最终结果见表6-5
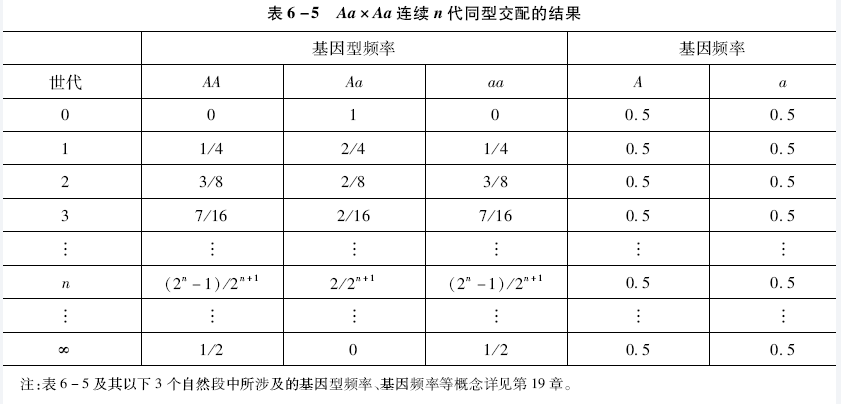
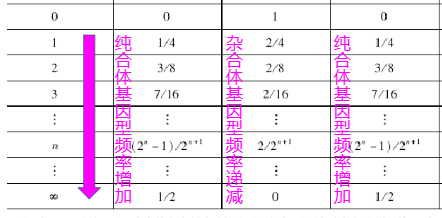
表6-5中结果显示
- 群体中Aa的基因型频率按Hn = 1/2H(n-1)(这里的n-1为下标) 的规律每代减少前一代的1/2而迅速递减,其极限值为0
- 同型交配群体中纯合体基因型频率每代都有增加,其频率各占1/2
- 完全同型交配的群体中的基因频率并没有改变

近交效应:
- 增加后代群体的纯和度,使遗传性状稳定.
亲缘关系越进的个体近交,其后代纯和体比率就上升越快,如每代进行同一类型的近交,自交6代后纯合子的比率可达成100%.
- 衰退现象的出现
由于纯合子比率增加,使原来处于杂合状态被掩盖的隐性形状得以表现,表现为生活力,繁殖力下降.抗病力下降等,如植物白化的出现.

(2) 近交系数与亲缘系数
- 近交系数(coeficient ofinbreeding,F或f) :个体在一个特定的基因座位上接受两个遗传上或者说血缘上相同的等位基因的概率.
或:个体从其双亲共同祖先得到一对遗传上是等同的纯合基因的概率,它反映了亲缘关系相近程度,是度量个体近交程度的重要遗传参数.
-
- 若某个体的F值大,说明其父母亲缘关系近
- 若F = 0,说明其双亲无亲缘关系
- 亲缘系数(coefficient of relationship ,R):有亲缘关系的两个个体的基因组中携带相同而且同源基因的比例。
它是两个个体亲缘程度的度量。亲缘系数愈大,亲缘关系愈近亲缘系数为0,则可认为个体(X、Y)间在近期几个世代内没有共同祖先。

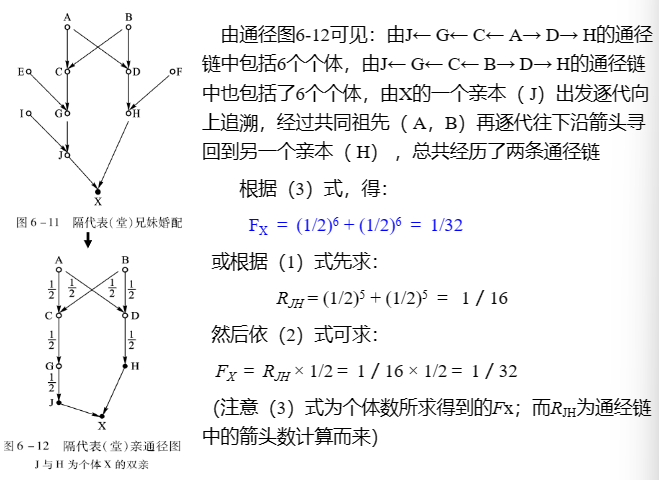
(3)运用通径分析方法计算近交系数和亲缘系数
Wright(1921)提出通径分析 ( path analysis)原理与方法
① 通径与通径链

在一个相关变量的网络系统中,连接“原因”与“结果”的每一个单箭头线条称为一条通径(path)
因果关系:单箭头线,方向由“因”到“果”,称为通径线
通径线的系数称为通径系数: 每一条通径中的原因”对于某一个“结果”所起作用的相对大小,用通径系数 (path coefficient ) 表示。即度量各“原因变量”对“结果变量”的直接影响的系数称为通径系数。
由一条或一条以上的通径所组成的完整的通道称为通径链。

② 通径分析的理论及其应用
通径分析证明,在随机交配群体中个体世代的每条通径的通径系数=1/2。而各类亲属关系的特定个体X、Y间的亲缘系数:👇

其中Rxy表示X、Y个体的亲缘系数,L表示沿着某两个特定亲属间(XY间)的连接通径链条中的箭头数,这个横过来的M表示所有这类链条之和。
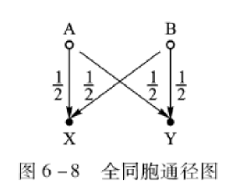
在图6-8通径图中由X←A→Y及由 X←B→Y 两条通径链连接X、Y两个特定个体,一条经过共同祖先A,另一条经过共同祖先B,而A、B两个个体不存在共同祖先,属随机婚配。每条通径链的系数由各通径的系数相乘,然后对每条通径链的系数求和则有:
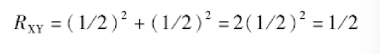



(4)近交降低群体基因型值的平均值,杂交提高群体均值
一个数量性状的基因型值是由基因的累加效应值和非累加效应值组成.
非累加效应值中的显性效应和大部分上位效应都存在于杂合体中,因此也可大致称非加性效应值为杂合效应值
- 上位效应 epistatic effect 定义:影响同一性状的两对非等位基因,其中一对基因 (显性或隐性的)抑制 (或掩盖)另一对显性基因的作用时所表现的遗传效应。
(5)近交使群体分化,杂交使群体一致
在一对等位基因(A,a)遗传的群体中,同型交配的最终结果是使群体分化成两个纯系,AA系与aa系。在两对基因 (A,a及B,b)的情况下,分化成AABB、AAbb、aaBB和aabb4 个不同的纯系。杂交则相反,它能使个体的基因型杂合化,因而使群体趋向一致。最为典型的情况是两个纯系AA与aa间的杂交,F群体中100%为Aa的杂合体,群体达到了完全的一致性.
(6)近交与人工选择相结合使提高杂种优势的重要手段之一
通过人工选择保留理想的纯合体,近交与人工选择结合能较快地加大群体间基因频率的差异,因而也成为提高杂种优势的有力手段
杂种优势及其遗传理论
(1) 概念:
杂种优势(heterosis )是指两个遗传组成不同的亲本杂交产生的杂种一代(F1),在生长势、生活力、繁殖力、适应性以及产量和品质等性状上比双亲优越的现象
利用杂种第一代这种超亲现象,以获得更大的经济效益,称为杂种优势利用.
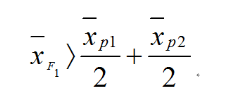
(2)杂种优势形成的机制有两种学说:
① 显性说(dominance hypothesis, Bruce, 1910)

② 超显性说(overdominance hypothesis)

该假说认为同基因座杂基因间及不同基因座之间,是复杂的互作关系而不是显隐性关系,杂种优势的形成来源于基因的互作,其中包括同基因座在杂合状态下基因之间的互作和不同基因之间的基因互作(亦称上位性互作)。在杂合状态下,基因表达水平可大大超过纯合状态的基因表达水平。由于基因型在杂合状态下的性状表现超过基因型在显性情况下的表现,故称为超显性现象.
超显性假说所说的杂种优势的生化基础两种可能情况
①两个等位基因各自编码一种蛋白质,这两种蛋白质的相互作用的结果比各自独立存在更有利于个体的生存。例如人类的镰刀形血红蛋白杂合体(HbA/HbS)的红细胞中同时存在着两种血红蛋白:成人血红蛋白(HbA)和刀形细胞血红蛋白 (HbS)。杂合体既不是贫血症患者,又较不易为疟原虫感染,因而在疟疾流行的地区更有利于生存
②两个杂合等位基因所编码的多结合成为活力高于相同亚基所形成的蛋白质。等位基因的这一相互作用形式至少曾经在粗糙脉抱菌的谷氨酸脱氢酶基因中发现
上述两种假说一个强调显性基因作用,另一个强调基因间相互作用,它们既非互相排斥又不能概括一切。根据数量性状的遗传分析杂种优势的遗传实质在于显性效应、累加效应以及异位显性、互补作用和超显性等各种基因互作效应。在某一材料中可能只有某一种基因互作决定某种程度的杂种优势
希望能帮到你~