Characterization and Functional Analysis of 4-Coumarate:CoA Ligase Genes in Mulberry
桑树中4-香豆酸:辅酶A连接酶基因的表征与功能分析
桑树T2T基因组-文献精读16

摘要
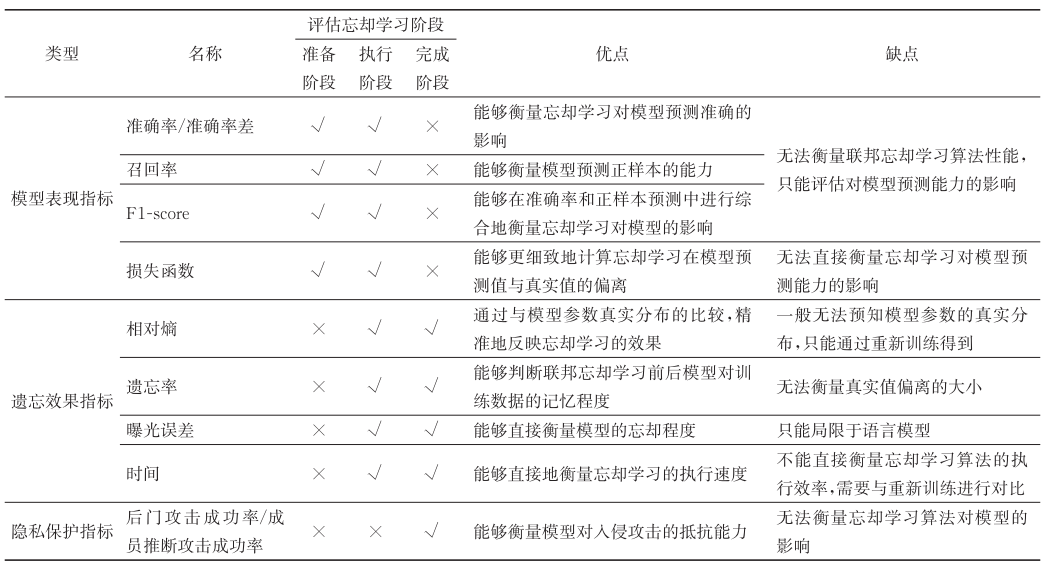
4-香豆酸:辅酶A连接酶(4CL)由一个小型的多基因家族编码,能够催化羟基肉桂酸类与辅酶A(CoA)的连接,这是代谢物进入类黄酮或单木质素途径的分支点。在本研究中,我们从M. notabilis基因组数据库中鉴定了四个4CL基因,并从紫梢木(M. atropurpurea cv. Jialing No.40)中克隆了四个Ma4CL基因。组织特异性表达分析表明,Ma4CL3的表达水平高于其他基因,且Ma4CL3在根皮、茎皮和老叶中的表达尤为强烈。此外,Ma4CL3的表达模式与果实发育过程中类黄酮总含量的变化趋势相似。系统发育分析表明,Mn4CL1、Mn4CL2和Mn4CL4属于I类4CL,而Mn4CL3属于II类4CL。不同的Ma4CL基因对一系列胁迫的反应各不相同。在创伤处理、水杨酸处理和紫外线处理后,Ma4CL3的表达水平高于其他Ma4CL基因。体外酶活性分析表明,在肉桂酸、4-香豆酸和咖啡酸之间,4-香豆酸是最佳底物,但对阿魏酸和愈创木酸没有催化活性。亚细胞定位实验结果表明,Ma4CL3定位于细胞膜,能够激活转录。我们采用不同的载体和策略,将Ma4CL3与白藜芦醇合酶(STS)融合,构建了四个Ma4CL-MaSTS共表达系统,以生成白藜芦醇。结果表明,只有使用T7启动子和乳糖操纵子表达MaSTS的转录融合载体pET-Ma4CL3-T-MaSTS能够合成白藜芦醇。
引言
从初级代谢向一系列苯丙烷类次生产物的合成转移碳流至少涉及三个酶促步骤,这些步骤分别由苯丙氨酸解氨酶(PAL)、肉桂酸4-羟化酶(C4H)和4-香豆酸:辅酶A连接酶(4CL:EC 6.2.1.12)催化。这些酶共同形成了通用的苯丙烷类代谢途径,保持苯丙烷类化合物合成的连续代谢流动【1,2】(图1)。4CL是最后的关键酶,催化4-肉桂酸或其衍生物转化为相应的辅酶A酯,生成所需的前体物质,这些前体用于多种天然产物的形成,如类黄酮、白藜芦醇和木质素,这些物质具有多样化的功能,如作为植物抗真菌感染的植保素、紫外线防护剂、花卉和果实的色素以及细胞壁的结构成分【3-5】。

图1. 通用苯丙烷类代谢途径中的酶由黑色箭头连接,包括PAL、C4H和4CL。
虚线箭头表示从通用苯丙烷类代谢途径发出的木质素生物合成支路,该支路同样由4CL催化。灰色箭头表示查尔酮和白藜芦醇生物合成的途径。
4CL基因在植物中以多基因家族的形式存在,包括拟南芥(四个成员【6】)、水稻(五个成员【7】)、三裂杨(四个成员【8】)和藓类植物(四个成员【9】)。然而,4CLs在许多植物中可分为两类:I类和II类。I类与木质素的生产关系更为密切,而II类4CL更可能参与类黄酮的生物合成【6,10,11】。先前关于木质素合成的研究表明,抑制4CL表达可使烟草、拟南芥、辐射松和颤杨等植物的木质素生产减少25%至45%【10,12-14】。此外,苯丙氨酸代谢途径的一个分支产物——白藜芦醇,已被证明对人类健康有益【15-17】。近年来,研究还探讨了通过4CL与其他基因共表达,在发酵过程中合成非天然类黄酮、白藜芦醇及其他具有生物活性的物质【18,19】。
桑树(Morus alba L.)的叶片是家蚕(Bombyx mori)的最佳食物。人类对桑树-蚕互动的利用至少可以追溯到5000年前,且对世界历史产生了巨大影响【20】。此外,由于其丰富的次生代谢产物,桑树的不同部位因其多种健康益处而被广泛研究。桑葚被广泛认为是一种营养丰富的食物,其花青素含量是所有水果中最高的之一【21】。此外,桑树的叶、根皮和枝条长期以来在中药中使用【22】。近期研究表明,桑树中的一些苯丙烷类代谢产物(如花青素、白藜芦醇和桑黄素A)对人类健康具有抗氧化、抗高血糖、降脂和抗动脉粥样硬化等作用【15-17,23】。然而,负责合成这些具有生物活性的化合物的基因在桑树中尚未被明确。因此,有必要研究Ma4CL基因在桑树中的功能。本研究报告了从桑树中分离和表征四个Ma4CL基因,确定其组织特异性表达,监测其对创伤、紫外线、水杨酸(SA)、NaCl和脱落酸(ABA)胁迫的响应,并表征了Ma4CL3的亚细胞定位。此外,我们评估了不同的策略,将Ma4CL基因与白藜芦醇合酶(STS)融合,构建了四个Ma4CL-MaSTS共表达系统,以生成白藜芦醇。
材料与方法
植物材料与多重胁迫测试
桑果材料采集自西南大学桑树园的桑树品种“嘉陵40号”的七个不同发育阶段。1周龄的黄化幼苗用于ABA(100μg/L)和NaCl(50mg/L)实验,在25°C下16小时光照、8小时黑暗的周期中培养,分别在1、3、6、10、16和24小时后采集。此外,采集了10周龄桑树叶片,用于紫外线-C(254 nm, Spectroline model ZW30S18Y,输出功率110 μW/cm²,距离1米)、SA(100μg/L)和机械创伤处理。在SA处理中,将SA溶液喷洒在叶片上,机械创伤通过使用无菌牙签沿叶脉制造八个伤口完成【24】。在紫外线处理中,植物在距离15厘米的范围内接受紫外线-C照射。所有材料都被冷冻在液氮中,并储存在−80°C中。
数据检索与Ma4CL cDNAs的克隆
利用毛白杨(Populus tomentosa,AFC89538.1)、陆地棉(Gossypium hirsutum,ACT32027.1)和拟南芥(Arabidopsis thaliana,NP_175579.1)基因编码区在桑树基因组数据库(MorusDB - Mulberry Genome Database)中进行相似性搜索。使用BLASTN和SMART工具(SMART: Main page)筛选候选基因。最终选择了四个Ma4CL基因进行进一步研究。使用引物(见表S1)从“嘉陵40号”中分离Ma4CL基因,纯化后的PCR产物通过测序确认(见文件S1)。
序列比对与系统发育分析
使用MEGA 5.0软件进行多序列比对,并使用MUSCLE 3.6软件基于最大似然法构建系统发育树。系统发育重建所需的其他4CL氨基酸序列从Genbank数据库中获取,包括毛白杨(Poptr4CL1-5 3A9U_A, AFC89538.1, AFC89539.1, AFC89540.1, AFC89541.1)、陆地棉(Gh4CL1 ACT32027.1)、烟草(Ntab4CL2 O24146.1)、黄松(Pinta4CL P41636.1)、拟南芥(At4CL1-4 NP_175579.1, NP_188761.1, NP_176686.1, NP_188760.3)、稻(Os4CL1-5 NP_001061353.1, NP_001047819.1, NP_001046069.1, NP_001058252.1, NP_001061935.1)、多年生黑麦草(Lp4CL1 AAF37732.1)、紫草(Le4CL1-2 BAA08365.1, BAA08366.2)、覆盆子(Ri4CL1-3 AAF91310.1, AAF91309.1, AAF91308.1)和大豆(Gm4CL1-4 NP_001236418.1, NP_001236236.1, NP_001237270.1, P31687.2)。
RNA提取、cDNA合成及定量反转录-聚合酶链反应(qRT-PCR)
从根皮、茎皮、老叶和嫩叶中提取总RNA,使用RNA提取试剂盒(TaKaRa,日本滋贺),桑果总RNA提取使用RNA提取试剂盒TransZol Plant Kit(TransGen Biotech,北京)。为消化基因组DNA,RNA样品经DNase I(TaKaRa)处理,使用2 μg纯化的RNA按生产商说明用Moloney鼠白血病病毒反转录酶(Promega,美国威斯康星州麦迪逊)合成cDNA。使用七倍稀释的cDNA进行qRT-PCR,引物使用GeneScript公司的在线工具设计【25】。通过SYBR® Premix Ex TaqTM II(TaKaRa)按生产商的说明制备反应,2 μL稀释后的cDNA用于20 μL的反应中。qRT-PCR在StepOne实时PCR系统
(Applied Biosystems,美国加州福斯特城)中进行。使用MaACTIN3(HQ163775.1)作为内参基因,以标准化目标基因的相对表达量【25】。所有数据使用2−ΔΔCt法分析。
总类黄酮含量测定
总类黄酮含量测定按照之前描述的方法进行【26】。收集不同发育阶段的桑果,使用真空冷冻干燥机(Thermo Fisher Scientific,美国马萨诸塞州沃尔瑟姆)将其干燥。每个样品(0.5 g)在低温下磨成粉末,三次用10 mL 75%(v/v)乙醇提取1小时,滤液蒸干后溶解在5 mL 75%(v/v)乙醇中。随后,向0.1 mL提取液中加入0.5 mL NaNO2(50 mg/mL),剧烈摇匀后室温放置6分钟。然后向反应混合物中加入0.5 mL Al(NO3)3。室温放置6分钟后,加入2.5 mL NaOH(1M)混合并在室温放置15分钟。吸光度在510 nm处测定。
Ma4CL3蛋白的亚细胞定位
将Ma4CL3分别与增强型绿色荧光蛋白(EGFP)基因的5'和3'末端融合,以构建融合蛋白表达盒。首先将EGFP插入植物表达载体pLGNL的Spe I/Bam HI位点,构建pLGNL-EGFP。然后将Ma4CL3插入BamH I/Kpn I和EcoR I/Spe I位点,分别构建pLGNL-Ma4CL3::EGFP和pLGNL-EGFP::Ma4CL3。包含EGFP、Ma4CL3::EGFP和EGFP::Ma4CL3融合基因的载体通过测序确认(Sangon Biotech,上海),然后通过农杆菌LBA4404瞬时转化洋葱表皮细胞。染色后使用荧光显微镜拍摄图像。
重组蛋白表达与纯化
全长的Ma4CL3 cDNA被克隆到pET28a(+)载体的Hind III和Xho I位点之间(见表S3),以获得包含氨基末端六组氨酸标签的融合蛋白。所得质粒转化到Escherichia coli BL21 (DE3) pLysS细胞中培养,培养至600 nm的光密度(OD600)达到约0.6时,加入0.1 mM异丙基-β-D-硫代半乳糖苷(IPTG)进行诱导,16°C下培养12小时,然后通过离心收集细胞。重组酶通过Ni-NTA金属亲和基质(GenScript)根据生产商的说明进行纯化。然后在12%的聚丙烯酰胺凝胶的泳道中识别蛋白条带,使用UltraFlex III基质辅助激光解吸电离-飞行时间串联质谱(MALDI-TOF/TOF MS,见文件S2)进行分析。从MALDI-TOF/TOF MS获得的肽片段离子数据用于在桑树基因组数据库(MorusDB - Mulberry Genome Database)中搜索蛋白候选物。
酶活性测定
酶活性测定按照之前描述的方法【6】进行,并进行了少量修改。反应体系包括2 mL 100 mM Tris-HCl缓冲液,包含5 μg亲和纯化的蛋白、2.5 mM MgCl2、2.5 mM ATP,以及一系列的底物,包括肉桂酸、4-香豆酸、咖啡酸、阿魏酸和愈创木酸。通过加入0.2 mM CoA开始反应。分别在311 nm、333 nm、345 nm、346 nm和352 nm处测量相应的肉桂酰辅酶A、4-香豆酰辅酶A、阿魏酰辅酶A、咖啡酰辅酶A和愈创木酰辅酶A产物的吸光度,对比空白组。相应的CoA酯复合物的消光系数(ε)分别为22、21、19、18和20 mM−1 cm−1。
发酵实验
为了研究Ma4CL3的其他特性,使用了多种策略将其与MaSTS(GenBank: ALS20364.1)共同表达。所有用于生成融合蛋白的引物列于表S3,融合策略如图2所示。

图2. 表示Ma4CL3和MaSTS蛋白共表达的示意图
4-S由pETDuet-1/pACYCDuet-1质粒携带;S4和4S由pET28a(+)质粒携带;4-T-S也由pET28a(+)质粒携带,但有独立的T7启动子和乳糖操纵子。
4-S
Ma4CL3基因通过引物pETDuet4CL3-F(包含Pst I位点)和pETDuet4CL3-R(包含Hind III位点)扩增,并克隆到pETDuet-1质粒的相应位点,生成pETDuet-4CL3载体。MaSTS基因通过引物MaSTS-F(Kpn I位点)和MaSTS-R(Xho I位点)扩增,并插入pETDuet-4CL3载体的Xho I/Kpn I位点,生成pETDuet-4CL3-STS质粒。pACYCDuet-4CL3-STS质粒按相同方法组装。
S4和4S
根据之前的研究【27】,我们还生成了两种不同的Ma4CL3和MaSTS融合蛋白,携带于pET28a(+)质粒。第一种是Ma4CL3与MaSTS的氨基末端融合,生成pET4CL3-STS,称为4S。第二种是MaSTS与Ma4CL3的氨基末端融合,生成pETSTS-4CL3,称为S4。最靠近启动子的第一个基因的终止密码子被突变为TCA(编码Ser),并在第二个基因前插入了一个迷你连接肽,编码Ser-Ser-Gly-Ser-Gly。
4-T-S
Ma4CL3基因在5'端携带BamH I位点,3'端携带Kpn I和Xho I位点,首先将其克隆到pET28a(+)载体的BamH I和Xho I位点,生成pET4CL3载体。pET28a(+)片段从AGATCTCGATCCCGCGAA突变为CTGCAG(Kpn I位点),并通过融合PCR将其连接至MaSTS的5'端。然后,将消化后的融合片段插入pET4CL3的Kpn I/Xho I位点,生成pET4CL3-T-STS。因此,Ma4CL3和MaSTS基因位于T7启动子/乳糖操纵子和核糖体结合位点之前。
上述四个共表达质粒分别转化至E. coli BL21 (DE3) pLysS中。细胞在28°C培养至OD600值达到0.6时,加入0.1 mM IPTG进行诱导,在16°C下培养10小时以生产蛋白。然后通过5,000 × g离心5分钟收集细胞,重悬于含1 mM 4-香豆酸、0.1 mM马洛酰辅酶A、0.1 mM IPTG和必要抗生素的M9培养基中。细胞在25°C下孵育24小时后,超声破碎发酵液,并通过12,000 × g离心20分钟沉淀不可溶碎片。随后用乙酸乙酯提取,收集1 mL有机相样品进行高效液相色谱(HPLC)分析。
HPLC分析
分析采用Waters 2487 HPLC系统(美国马萨诸塞州米尔福德)和5 μm C18反相柱(4.6×150 mm,Waters SunfireTM,爱尔兰都柏林)。注射体积为10 μL,流速为0.8 mL/分钟。分离温度为25°C,检测波长为306 nm。乙酸乙酯样品离心后通过0.22 μm滤膜过滤。流动相为甲醇(A)和0.1%乙酸水溶液(B)。溶剂梯度洗脱程序如下:0–10分钟,10–30%A,90–70%B;10–25分钟,30–50%A,70–50%B;25–30分钟,50–70%A,50–30%B;30–45分钟,70–30%A,30–70%B;45–50分钟,30–10%A,70–90%B。
统计分析
所有qRT-PCR实验均进行三次重复,结果以均值±标准差(SD)表示。使用SPSS Statistics 18.0(美国伊利诺伊州芝加哥)进行Duncan多重范围检验的统计分析,图表使用GraphPad Prism v5.0软件绘制(GraphPad Software Inc.,美国加州拉霍亚)。处理之间均值的显著性差异用“*”表示。
结果
4CL基因的结构和系统发育分析 根据桑树基因组数据库和GenBank获取的序列进行多序列比对,发现了四个Mn4CL基因,并通过分离各种组织的对应cDNA确认了这些基因。与其他物种中的4CL基因类似,Mn4CL基因具有多个外显子和内含子结构。Mn4CL1和Mn4CL3含有六个外显子和五个内含子,Mn4CL2含有五个外显子和四个内含子,而Mn4CL4仅含有两个外显子和一个内含子。所有Mn4CL基因均预测编码家族蛋白,Mn4CL1含有646个氨基酸,Mn4CL2含有546个氨基酸,Mn4CL3含有595个氨基酸,Mn4CL4含有407个氨基酸。系统发育分析结果将这些序列明显划分为两组,表明Mn4CL3属于II类,而Mn4CL1、Mn4CL2和Mn4CL4属于I类。多序列比对表明这些桑树蛋白中的催化结构域和活性位点是保守的,四个Mn4CL都具有两个结构域,即假定的AMP结合基序(box I)和保守的box II域。

图3. Morus notabilis 4CL基因的基因结构
模式图显示了外显子(灰色框)和内含子(外显子之间的虚线)。基因结构由Fancy Gene工具展示(http://bio.ieo.eu/fancygene/)。

图4. Mn4CL蛋白的系统发育树分析
系统发育分析结果将这些序列分为两组,表明Mn4CL3属于II类,而Mn4CL1、Mn4CL2和Mn4CL4属于I类。

图5. 推导出的氨基酸序列的比较
Box I 代表推测的 AMP 结合域,Box II 代表保守的 GEICIGR 推测催化位点。
不同组织中 Ma4CL 基因的表达及其在多重胁迫下的动态表达
为了进一步研究 Ma4CL 基因在各种组织中的表达情况,通过使用特异性引物(S2 表)进行 qRT-PCR 定量测量了四个 Ma4CL 基因的转录水平。从10周龄的桑树植物的根皮、茎皮、老叶和嫩叶中分离了总 RNA。如图 6A 所示,所有的 Ma4CL 基因在所有选定的器官中均有表达,但它们的表达量存在显著差异。Ma4CL3 的表达水平高于其他三个基因,且 Ma4CL3 在根皮、茎皮和老叶中强烈表达。此外,Ma4CL3 在老叶中的转录水平最高,其次是根皮和茎皮,而在嫩叶中的转录水平最低。此外,Ma4CL1 和 Ma4CL2 在根皮中高度表达,而 Ma4CL4 在所有选定的器官中的表达量最低。

图6. Ma4CL基因在不同组织中的表达及在多种胁迫下的不同响应模式
所有的Ma4CL基因在所有选定的器官中都有组成型表达(a),但其表达量存在显著差异。Ma4CL3在相同组织中的表达水平高于其他基因,并且在根皮、茎皮和老叶中强烈表达。在多种胁迫下,Ma4CL基因表现出不同的响应模式,其中Ma4CL3在创伤(b)、水杨酸(SA)(c)和紫外线(UV)(d)处理后的表达诱导程度高于其他基因。每个柱状图上的误差条表示三次重复的标准差(SD)。
不同4CL基因的表达在受到创伤时被诱导(图6B)。Ma4CL1和Ma4CL3的表达呈现“M型”模式,在叶片受伤后1小时和24小时达到峰值。尤其是Ma4CL3在24小时时的诱导明显,其第二个表达峰值是对照组的4.11倍。然而,Ma4CL2和Ma4CL4对创伤处理的响应较弱,它们的转录水平分别在1小时和3小时达到峰值。
Ma4CL基因对SA处理表现出不同的响应(图6C)。Ma4CL3对SA敏感,在1小时时达到峰值,6小时时下降到正常水平,10小时后其表达急剧上升,此时其表达量约为对照组的7.06倍。Ma4CL1、Ma4CL2和Ma4CL4的表达模式相似,表现为SA处理后略有上升,随后回到对照水平。
Ma4CL基因的表达受到紫外线诱导(图6D)。除了Ma4CL4在6小时时表现出单峰表达外,其他Ma4CL基因表现出类似的双峰模式,并在10小时时下降。然而,Ma4CL1的表达在3小时和16小时达到峰值,而Ma4CL2在6小时和16小时达到峰值。与其他处理类似,Ma4CL3的表达在紫外线照射下显著诱导,并在1小时达到最大值,约为对照组的1.81倍。
Ma4CL基因表达及桑葚果实发育过程中的总黄酮含量测定
在桑葚果实发育的S1至S7阶段(图7E)中,四个Ma4CL基因在整个果实发育过程中不断表达,但它们表现出两种不同的表达模式(图7A-7D)。Ma4CL1、Ma4CL2和Ma4CL4在S1阶段表达较高,然后在S2阶段急剧下降,随后逐渐增加,分别在S6、S3和S3阶段达到最大水平,然后在后续果实发育中下降。然而,Ma4CL3的表达模式与其他基因明显不同;其转录水平在S4之前下降,然后迅速上升,在S5阶段达到峰值,随后缓慢下降(图7C)。在桑葚果实发育的S1至S7阶段,动态总黄酮含量也得到了测定。如图7E所示,总黄酮含量的趋势与Ma4CL3在果实发育中的表达模式相似,尽管在S6阶段达到了峰值。

图7. 桑椹果实发育过程中Ma4CL基因的表达及总黄酮含量
a、b、c 和 d 分别代表 Ma4CL1、Ma4CL2、Ma4CL3 和 Ma4CL4 的表达水平,e 和 f 分别表示果实发育阶段 S1 到 S7 期间的总黄酮含量。每个柱状图上的误差条表示三次重复的标准差(SD)。
在不同胁迫条件下,Ma4CL基因表现出不同的上调表达模式。在NaCl胁迫下(图8),Ma4CL1和Ma4CL3在茎中的表达在1小时时达到峰值,随后下降(图a和c)。Ma4CL2和Ma4CL4的表达表现出类似的双峰模式(图8b和8d)。在根中,Ma4CL1和Ma4CL2的表达分别在6小时和3小时达到峰值(图8e和8f)。然而,Ma4CL3和Ma4CL4的表达并未明显增加,并且在1小时和12小时明显下调(图8g和8h)。

图8. NaCl处理下Ma4CL基因的相对表达水平
Ma4CL1和Ma4CL3在茎中的表达在1小时达到峰值,Ma4CL2和Ma4CL4则表现出类似的双峰表达模式(图a–d)。在根部,Ma4CL1和Ma4CL2的表达分别在6小时和3小时达到峰值。然而,Ma4CL3和Ma4CL4的表达在1小时和12小时被下调(图e–h)。每个柱状图上的误差条表示三次重复的标准差(SD)。处理之间的显著差异(p < 0.05)用“*”标记。
在ABA处理下,茎和根表现出不同的反应(图9)。当茎受到ABA处理时,Ma4CL1和Ma4CL2的表达分别在1小时和12小时达到峰值,随后下降(图9a和9b),而Ma4CL3和Ma4CL4的表达水平并没有显著变化(图9c和9d)。然而,根中Ma4CL基因的表达表现出完全不同的反应。除了Ma4CL3在3小时达到最大表达(图9g)外,其他基因的表达水平逐渐增加,并在处理24小时后达到最大值(图9e、9f和9h)。此外,根和茎的表达水平比率(RERS)也表现出差异(表1)。从表1可以明显看出,Ma4CL2的RERS在24小时之前低于对照组,而Ma4CL3和Ma4CL4的RERS则高于对照组。

图9. ABA处理下Ma4CL基因的相对表达水平
Ma4CL基因在茎(a–d)和根(e–h)中表现出不同的响应。在茎中,Ma4CL1和Ma4CL2的表达分别在1小时和12小时达到峰值,而Ma4CL3和Ma4CL4的表达没有显著变化。然而,Ma4CL3在根中的表达在3小时后达到最大值,而其他基因的表达水平逐渐增加,并在24小时后达到最大值。处理之间的显著差异(p < 0.05)用“”标记。在ABA处理4天后(i),尽管所有的RERS(根茎表达比率)显著上调,Ma4CL3的表达仅上调了3.46倍,而Ma4CL1、Ma4CL2和Ma4CL4的表达则分别上调了约5.23倍、16.50倍和9.47倍,与对照组相比(“”表示p < 0.01)。(j) ABA处理下,根长与茎长的比值显著变化,主根比对照更粗壮,且根毛数量显著增加。每个柱状图上的误差条表示三次重复的标准差(SD)。

表1. ABA处理下根和茎的表达水平比率
经过4天的ABA处理,尽管所有的根茎表达比率(RERS)显著上调,Ma4CL3的表达仅增加了3.46倍,而Ma4CL1、Ma4CL2和Ma4CL4的表达分别上调了约5.23倍、16.50倍和9.47倍,与对照组相比(图9I)。此外,根长与茎长的比值达到了3.49,而对照组仅为1.12。此外,ABA处理组的主根比对照组更加粗壮,且根毛数量显著增加(图9J)。
Ma4CL3蛋白的亚细胞定位
为了研究Ma4CL3蛋白的定位,EGFP、Ma4CL3::EGFP和EGFP::Ma4CL3融合基因在花椰菜花叶病毒(CaMV)35S启动子控制下(图10A),被转染到洋葱表皮细胞中。EGFP载体作为对照,并单独转染。可以清楚地观察到,EGFP的绿色荧光信号分布在细胞质、细胞核和细胞膜上(图10B)。然而,Ma4CL3::EGFP和EGFP::Ma4CL3融合蛋白的绿色荧光信号主要集中在细胞膜上,可能在该处激活转录,暗示Ma4CL3是一个定位于细胞膜的蛋白质。该结果与4CL家族蛋白的功能一致。

图10. Ma4CL3蛋白的亚细胞定位
(a) EGFP、Ma4CL3::EGFP 和 EGFP::Ma4CL3 的融合策略。(b) 阳性对照是仅包含 EGFP 的转染质粒。DAPI 用于指示细胞核。观察到 Ma4CL3 主要定位于细胞膜。
Ma4CL3的酶活性测定
Ma4CL3的开放阅读框被亚克隆到表达载体pET28a(+)中,并获得了可溶性蛋白质。通过金属亲和色谱法纯化后,使用MALDI-TOF/TOF MS对纯化的Ma4CL3蛋白进行分析,结果表明纯化蛋白的肽段完全匹配预测的肽段(S2文件)。对Ma4CL3使用多种底物进行酶活性测定,包括肉桂酸、4-香豆酸、咖啡酸、阿魏酸和松柏酸。正如表2所示,4-香豆酸的Km为10.49 μM;然而,Vmax仅为4.4 nkat mg−1。这表明在肉桂酸、4-香豆酸和咖啡酸中,4-香豆酸是最佳底物,而在使用松柏酸和阿魏酸作为底物时没有检测到催化活性。

表2. Ma4CL3的动力学特性 链接到表格
发酵测试
正如方法部分所述,使用了四种共表达策略。不幸的是,只有 4-T-S 策略在低温(16°C)和低IPTG(0.1 mM)发酵环境中产生了两种可溶性蛋白质,但在发酵液中仅产生了 0.1268 mg/L 的白藜芦醇(图11)。我们未能通过 4-S、4S 和 S4 策略获得可溶的 Ma4CL3、Ma4CL3-MaSTS 或 MaSTS-Ma4CL3 蛋白质。

图11. 发酵测试和HPLC检测结果
(a) 在重组大肠杆菌中构建的白藜芦醇的生物合成途径。白藜芦醇的标准色谱图 (b) 和乙酸乙酯提取物的色谱图 (c)。峰1代表4-肉桂酸,峰2代表白藜芦醇。
链接到图11
讨论
苯丙氨酸代谢途径是植物生长过程中碳积累的重要代谢途径之一。4CL是该途径的关键酶,受发育调控并可被外部刺激(如病原体感染、诱导剂处理、机械损伤和紫外线照射)激活【2,4】。此外,4CL对各种肉桂酸衍生物底物(如肉桂酸、4-香豆酸、咖啡酸、紫苏酸和阿魏酸)表现出不同的催化活性,并具有明确的底物偏好【9,28-30】。这种偏好有利于代谢过程,因为它可以为下游过程提供适当比例的肉桂酰辅酶A混合物,并调节代谢物流向木质素或非木质素的合成。
与其他物种的4CL基因类似,Mn4CL基因包含多个内含子和外显子,它们在长度和核酸序列上有所不同。然而,Mn4CL家族的box I和box II区域是保守的肽序列。box I(SSGTTGLPKGV)是AMP结合域,box II(GEICVRS)是催化位点【29】(图5)。在桑果不同生长阶段,Ma4CL3的表达水平高于其他三个基因,且Ma4CL3的表达趋势与果实中总类黄酮含量趋势一致。Ma4CL3的最大表达并未发生在果实颜色最深的S7阶段,而是在S5阶段。这与烟草花中Nt4CL2的表达模式相似,Nt4CL2在无色花冠管中表现出高表达,且在花的第4阶段首次观察到高Nt4CL2转录水平,而在第6阶段花中显著下降【2】。此外,根据桑树的组织特异性表达分析,Ma4CL3转录本主要存在于根皮、茎皮和老叶中。这些区域富含类黄酮【26,31】。体外酶活性测试中发现,重组Ma4CL3对4-香豆酸的亲和力大于其他底物,且4-香豆酰辅酶A是类黄酮生物合成最常用的前体。先前的研究表明,4CL对类黄酮的Km通常低于对单木质素的Km【6,9,28,30,32】。系统发育树表明Mn4CL3属于II类。在各种植物中,单木质素的生物合成被认为是I类4CL的主要功能,而II类4CL的主要功能与类黄酮合成有关【6,10,11】。最近的一项研究证明,拟南芥中的At4CL3突变显著降低了类黄酮苷的含量,约为野生型植物的80%【33】。此外,沉默At4CL1和At4CL2(I类)基因降低了植株总木质素含量【34】。沉默Os4CL3(I类)基因的表达显著减少了木质素含量【7】。基于上述实验,推测Ma4CL3的主要功能是类黄酮生物合成。
次生代谢物的合成是植物在应对真菌、紫外线和过氧化氢等应激条件下的重要防御反应之一【3,4,35】。由于类黄酮强烈吸收紫外线,类黄酮的诱导合成可为植物提供紫外线保护【4】。芹菜细胞悬浮培养体系被广泛研究,表明其在紫外线照射下会合成类黄酮【3,35】。研究显示,紫外线照射上调了几种基因的表达,包括PAL、4CL和CHS,这些基因对类黄酮合成至关重要【3,35】。当植物遭受机械损伤时,伤口部位的细胞壁首先通过蛋白交联进行加固,防止脱水和病原体感染,随后需要苯丙烷衍生物来木质化和木栓化细胞壁【36】。在拟南芥中,At4CL1和At4CL2(I类)转录本在受伤后迅速积累,但At4CL3(II类)对损伤无反应【6】。与之相反,本研究发现伤害显著诱导了桑树叶片中Ma4CL3的表达,而Ma4CL2和Ma4CL4的反应较弱。这表明Ma4CL3对伤害处理更为敏感。类似的伤害处理在银桦树叶片中诱导了Bp4CL1(I类)和Bp4CL2(II类)的表达,而Bp4CL3和Bp4CL4(I类)的转录水平则未发生变化【37】。此外,伤害已被证明可减少潜在木质素合成抑制因子的表达【39,40】,这可能是Ma4CL3在伤害应激下表达上调的原因。
桑树作为生态和经济上重要的多年生木本植物,能在严重荒漠化地区生长,适应干旱、风沙和盐碱环境【41】。在植物免疫反应中,ABA和SA作为重要的信号分子已被证明能提高植物的抗病、抗盐和抗脱水能力【42,43】。然而,关于SA对4CL调控的影响研究较少。本研究结果表明,SA诱导了四个Ma4CL基因的表达,且Ma4CL3的表达显著上调(比对照组高出7.06倍),而其他三个基因的表达没有变化。这表明Ma4CL3是SA应答中最重要的参与者。干旱和盐分增加了细胞中活性氧浓度,刺激ABA在植物根系中的合成和积累,从而诱导植物产生一定的生理和生化反应,使其能够在逆境中生存【43】。本研究发现,ABA处理下,Ma4CL基因在根部和茎部的表达有所不同。在ABA刺激下,桑树增强了地下部分的生长,而抑制了地上部分的生长,从而抵抗环境胁迫。例如,这提高了桑树从地下深处吸水的能力,同时减少了地上器官的水分蒸发。植物根系的发育也与木质素及Ma4CL基因RERS的增加有关。然而,NaCl和ABA胁迫下,Ma4CL基因的表达模式有所不同,这表明桑树通过ABA非依赖信号通路响应盐胁迫。
在植物进化过程中,同一家族基因的功能可能具有冗余或差异性。本研究发现,四个Ma4CL基因对外界刺激的反应不同。有些基因对外界刺激更敏感,而有些则不敏感,部分基因表现出相同的反应模式,这意味着这些同工型可能具有相似的调控作用,并在功能上相互冗余。通过监测转录水平和/或启动子分析,4CL基因在拟南芥【6,38】、烟草【44】、水稻【7】和白杨【10】中得到了检测。结果表明,4CL基因的调控十分复杂,既扮演正调控作用,也扮演负调控作用【38】。不同的4CL家族成员在应对生物和非生物胁迫时表现出不同的表达模式【6,7】。
众所周知,可溶性蛋白是蛋白活性的前提条件。在本研究中,尽管使用了多种表达载体,但在高浓度的IPTG或高温条件下仍未能获得可溶性Ma4CL3蛋白。先前的研究使用了4CL-STS共表达系统来发酵白藜芦醇,并且发酵液中的白藜芦醇含量远高于本研究【18,45–48】。需要指出的是,重组微生物生产白藜芦醇的效率取决于多种因素,如物种和菌株、转移基因的来源,以及使用的质粒或前体等其他参数【48】。例如,由BW27784(大肠杆菌)产生的白藜芦醇产量是烟草4CL(Nicotiana tabacum)和葡萄STS(Vitis vinifera)在酵母(Saccharomyces cerevisiae)中产量的2.67倍【45】。然而,拟南芥4CL(Arabidopsis thaliana)和花生STS(Arachis hypogaea)的白藜芦醇产量比烟草4CL和葡萄STS高出6.25倍【45,46】。同时,利用紫草4CL(Lithospermum erythrohizon)和花生STS的融合蛋白在酵母菌株中获得的白藜芦醇产量达到了5.25 mg/L【47】。在本研究中,我们探索了通过桑树基因发酵获得白藜芦醇的可能性。未来的研究将重点优化发酵条件,以获得更高浓度的白藜芦醇。
随着人们对健康问题的日益关注,天然产品越来越受到欢迎。然而,由于化学合成药物的副产物众多以及天然产物含量较低,广泛使用原材料对环境造成的破坏和污染也日益严重【26】。此外,桑树富含白藜芦醇、氧白藜芦醇和桑椹苷【31,49】,这表明桑树中存在一个高效的生物合成系统。我们发现Ma4CL的表达趋势与类黄酮合成相关,并建立了在微生物中共表达白藜芦醇的系统,作为从植物中提取或通过化学合成获取白藜芦醇的替代方法。这项研究为未来桑树苯丙氨酸代谢途径的研究奠定了基础。
结论
综上所述,本研究鉴定了来自桑树的四个Ma4CL基因。尽管这些基因在所有选定的器官中均有表达,但其表达存在显著差异。尤其是Ma4CL3在根皮、茎皮和老叶中的表达相对较高。基于体外酶活性分析、桑果中的表达以及对多种胁迫的响应,Ma4CL3的功能可能与类黄酮合成相关,且Ma4CL3定位于细胞膜。此外,我们建立了Ma4CL3和MaSTS的共表达体系,并通过发酵成功产生了白藜芦醇。这些结果为未来桑树苯丙氨酸代谢途径的研究铺平了道路。
小小 NCBI词条GenBank: AHL83551.1~!=