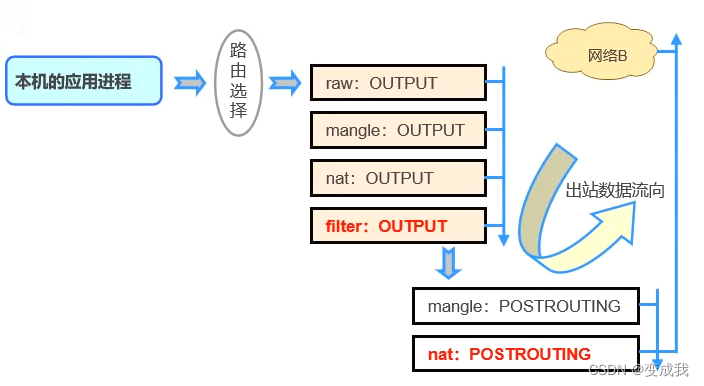
文献介绍

文献题目: Differences in neurotropism and neurotoxicity among retrograde viral tracers
研究团队: 曹罡(华中农业大学)、戴金霞(华中农业大学)
发表时间: 2019-02-08
发表期刊: Molecular Neurodegeneration
影响因子: 14.9
DOI: 10.1186/s13024-019-0308-6
摘要
Background: 基于嗜神经病毒的示踪剂已广泛应用于神经环路的绘制和操作。然而,它们的神经嗜性和神经毒性仍有待充分确定。
Methods: 通过神经环路示踪,我们系统地比较了不同的跨多突触(multi-trans-synaptic)和单突触(mono-synaptic)逆行病毒示踪剂,包括伪狂犬病毒(PRV)、狂犬病毒(RV)和新设计的逆行腺相关病毒(rAAV2-retro)示踪剂之间的神经嗜性差异。利用 scRNAseq 分析寻找神经嗜性差异的原因,并比较由糖蛋白缺失的 RV(RV-ΔG)和 rAAV2-retro 病毒感染引起的细胞毒性。采用免疫组化方法评价病毒毒性诱导的小胶质细胞活化和神经元蛋白的变化。
Results: 跨多突触逆行病毒示踪剂 PRV 和 RV 在用于从腘淋巴结示踪中枢神经环路时,表现出不同的神经嗜性。单突触逆行示踪剂,包括 RV-ΔG 和 rAAV2-retro,在应用于示踪下丘脑外侧区和内侧视前核的输入时,表现出不同的神经嗜性。rAAV2-retro 在大脑皮层中表现出偏好,而 RV-ΔG 更倾向于标记基底神经节和下丘脑。值得注意的是,当将 rAAV2-retro 注射到背外侧膝状核来标记初级视觉皮层时,我们发现它们对第 5 层和第 6 层的特异性丘脑神经层有明显的偏好。在对 RV 抵抗性神经元进行 TVA 受体基因的补充后,使得 EnvA-pseudotyped RV 感染成为可能,从而支持受体与病毒神经嗜性的关联。此外,无论是 RV-ΔG 还是 rAAV2-retro 在注射部位和逆行标记部位均表现出神经毒性影响,但 RV-ΔG 感染的变化更为显著。最后,我们证明了通过结合 rAAV2-retro、RV 和 rAAV 示踪剂,可以实现对特定目标核心进行更全面的高阶环路示踪的概念验证策略。
Conclusions: 不同的跨多突触和单突触逆行病毒示踪剂在大脑的某些区域表现出不同的神经嗜性,甚至皮层偏好。与 rAAV2-retro 相比,RV-ΔG 感染下观察到更多的神经毒性。通过结合 rAAV2-retro、RV 和 rAAV 示踪剂,可以实现高阶环路示踪。我们的研究结果为适当应用病毒示踪剂来描绘景观和解剖神经网络的功能提供了重要的参考。
Keywords: Neurotropic Virus, Retrograde tracing, pseudorabies virus (PRV), rabies virus (RV), rAAV2-retro, Neurotropism, Neurotoxicity, Multi-trans-synaptic Tracing, Mono-synaptic Tracing, RNA Sequencing
背景
人脑是一个高度复杂的网络,其中大量( )具有不同细胞类型的神经元连接形成大约 个突触。这种复杂的神经环路组织为认知、情感、记忆、感觉和运动等大脑功能奠定了结构基础。描绘神经环路的精细架构和功能对于理解复杂的大脑功能至关重要,并且在很大程度上取决于有效示踪和操纵技术的可用性。
在早期的神经环路研究中,几种示踪染料被用来标记突触前和突触后神经元,这有助于解决不同脑核之间的介观连接。辣根过氧化物酶 (HRP) 是第一个用于环路绘制的示踪剂,随后出现了一系列顺行或逆行示踪剂,包括碳菁染料(carbocyanine dyes)、羟芪巴脒(FluoroGold)、霍乱毒素(cholera toxin)、荧光微球(fluorescent microspheres)。然而,这些传统的神经示踪剂缺乏选择性,几乎没有跨突触能力,并且不能用于传递外源基因。
目前,基于嗜神经病毒的示踪剂最广泛用于识别突触前或突触后神经元、高阶神经连接、神经环路功能。不同的病毒,例如腺相关病毒(AAV)、狂犬病病毒(RV)、伪狂犬病病毒(PRV)、犬腺病毒(CAV)、单纯疱疹病毒(HSV)已被设计用于追踪和操纵神经环路。值得注意的是,跨单突触示踪工具的发展使得能够研究某些神经环路内的直接上游连接。最近,一种工程 AAV 变体 rAAV2-retro 被检查具有逆行追踪潜力,尽管其作用机制仍知之甚少。
相对于传统的示踪剂,嗜神经病毒由于病毒基因组复制和受体神经元中的外源基因表达而产生更强的信号。此外,这些示踪工具可以借助特定启动子或 Cre/lox 技术选择性地标记特定神经元。此外,表达光遗传学、化学遗传学、Ca2+/电压敏感基因的嗜神经病毒可用于监测和操纵特定神经环路内的活动。同时,之前的研究表明,天然的和工程的神经病毒都表现出对标记方向的偏好。单纯疱疹病毒 HSV 129 株主要以顺行方向运输。水泡性口炎病毒(VSV)糖蛋白包膜的 RV 也被用作顺行示踪剂。而 RV、CAV、缺乏 gE 和 gI 基因的 PRV 和 rAAV2-retro 病毒表现出对逆行标记的显着偏好。鉴于这些优点,基于神经病毒的追踪工具非常适合绘制精细神经环路及其功能。
尽管这些顺行和逆行嗜神经病毒极大地促进了神经环路连接和功能的解析,但值得注意的是,不同的示踪病毒可能会导致标记结果的差异。也就是说,虽然某些环路可以被不同的病毒同等地追踪,但对于不同的病毒将存在对其他环路的选择性或优先追踪。据报道,HSV1 和 PRV 可以感染神经元,包括支配注射部位的初级感觉神经元、运动神经元、交感神经元、副交感神经元。然而,这两种病毒对小型初级感觉神经元和自主神经元的亲和力比对运动神经元的亲和力更大。由于当注射到灵长类动物、大鼠和豚鼠的肌肉中时,支配肌肉的自主神经和感觉神经元很少被 RV 株 CVS-11 感染,因此该病毒被提议作为这些动物运动神经元网络的特异性示踪剂。Albisetti et al. 确定了两类感觉神经元,它们对来自脊髓的直接和跨突触 RV 感染具有抵抗力,这可能是由于病毒吸附受损所致。有趣的是,据报道,新设计的 CVS-N2cΔG 和 RV 具有密码子优化的嵌合糖蛋白(G),其中 G 来自 B19G RV 菌株和 Pasteur RV 菌株,可以提高示踪效率,这表明即使是不同的 RV 菌株可能具有不同的神经嗜性。此外,当 CAV2-Cre 和 HSV1 被应用于从内侧前额叶皮层到基底外侧杏仁核的逆行追踪时,它们在基底外侧杏仁核中表现出不同的嗜性。这种独特的神经嗜性很可能归因于不同神经元中嗜神经病毒入侵所需的不同受体的表达。然而,不同示踪病毒的受体仍有待充分表征。尽管不同的病毒示踪剂广泛应用于神经环路研究,但很少有研究系统比较不同嗜神经病毒的示踪特性,极大地阻碍了其在阐明神经环路方面的正确应用。
此外,据报道,几种病毒示踪剂在环路追踪过程中会引起受感染神经元的细胞毒性。HSV1 和 PRV 会引起受感染神经元的快速神经元变性。对于糖蛋白缺失的狂犬病病毒,虽然在感染早期阶段细胞病变可以忽略不计,但注射后 16 天左右会导致电生理特性改变,甚至神经元死亡。然而,对不同环路示踪剂引起的神经毒性的系统比较分析尚未得到充分记录。
因此,在本研究中,我们的目的是系统地比较当前神经科学研究中广泛使用的不同逆行追踪病毒,包括 PRV、RV 和新设计的 rAAV2-retro 病毒的神经毒性差异。我们进一步旨在通过使用 EnvA-TVA 互补系统并通过 scRNAseq 分析不同病毒标记神经元组中的基因表达谱来阐明这些追踪病毒的神经毒性差异背后的机制。然后通过 RNA-seq 和免疫组织化学比较 RV-ΔG 和 rAAV2-retro 病毒示踪剂的神经毒性。最后,我们通过结合不同的病毒示踪剂展示了高阶环路追踪的概念验证策略。我们的研究结果可能为神经环路结构和功能的综合研究中正确应用嗜神经病毒提供重要参考。
研究结果
1. PRV 和 RV 在跨多突触逆行追踪过程中表现出神经嗜性差异
伪狂犬病病毒(PRV)是最常用于对来自外周神经支配目标的中枢神经环路进行跨多突触追踪(trans-multiple synapse)的病毒。在本研究中,我们比较了固定 RV 衍生物 RV-B2C-EGFP(EGFP 插入 RV-B2C 菌株的 G- 和 L- 编码序列之间)与 PRV-Bartha 衍生物 PRV-152 的神经毒性作用(Fig. 1a)。由于腘淋巴结在解剖学上是孤立的,并且大小适合精确的病毒注射,因此选择它用于跨多突触逆行示踪病毒的神经性比较。这些表达 GFP 的病毒被注射到小鼠左腘淋巴结中,以分别追踪中枢神经系统的上游跨多突触环路。
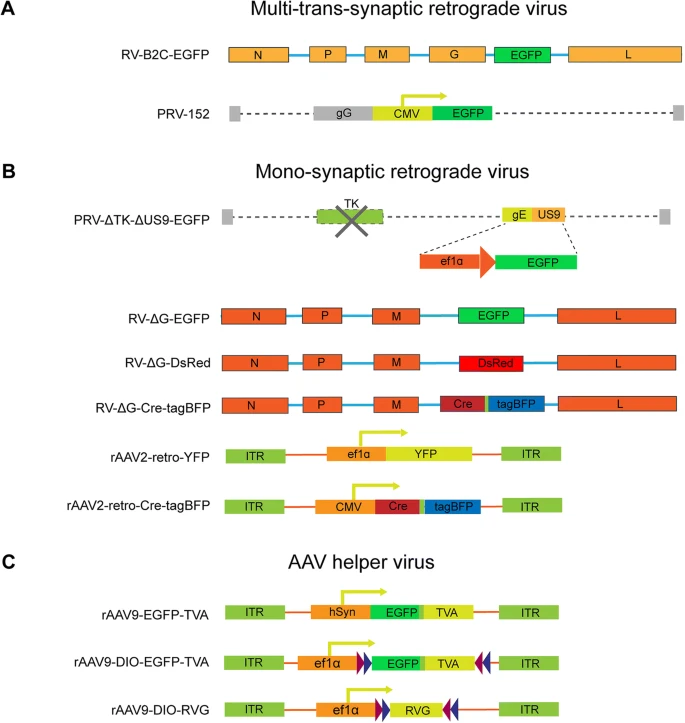
(a) 跨多突触逆行病毒的结构说明,包括 RV-B2C-EGFP 和 PRV-152。对于 RV-B2C-EGFP,EGFP 基因插入 CVS-B2C 菌株的 G- 和 L- 编码序列之间。对于 PRV-152,EGFP 由 Bartha 菌株的 gG 基因之后的 CMV 启动子驱动。
(b) 说明了单突触逆行病毒的结构。由于 TK(胸苷激酶)基因参与病毒复制,而 US9 负责顺行运输,因此通过删除 TK 和 US9 基因获得逆行单突触示踪病毒 PRV-ΔTK-ΔUS9-EGFP,其中位置 US9 基因通过同源重组在 Becker 菌株的 BAC 骨架上被 CMV 启动子和 EGFP 基因取代。对于 RV-ΔG 单突触病毒,将 EGFP、DsRed、Cre-T2A-tagBFP 分别插入狂犬病毒基因组中以替换 G 基因。对于rAAV2-retro,ef1a 和 CMV 启动子分别用于驱动 YFP 和 Cre-T2A-tagBFP 表达。
(c) AAV 辅助病毒的结构。hSyn 和 ef1a 启动子用于驱动 TVA、DIO-TVA 和 DIO-RVG 元件。
几个大脑区域,包括初级运动皮层(M1)、初级体感皮层(S1)、外侧导水管周围灰质(LPAG)(Fig. 2a, c, e, g)、视前区(VMPO, MPA, MPOL+MPOM, LPO)和终纹床核(BNST)(Fig. 2j)均被两种病毒示踪剂标记。然而,一些大脑区域被 PRV-152 或 RV-B2C-EGFP 专门或优先标记。将 RV-B2C-EGFP 注射到腘窝淋巴结时,丘脑腹侧后内侧核(VPM)、小脑(Fig. 2b and d)、纹状体(CPu)和次级躯体感觉皮层(S2)(Fig. 2j)观察到逆行标记,而注射 PRV-152 时没有观察到逆行标记。相反,室旁核(PVN)、蓝斑(LC)、巴林顿核(Bar)(Fig. 2f and h)、桥脑网状核(PnO)和巨细胞网状核(Gi)(Fig. 2j)中有大量神经元大部分由 PRV-152 标记。冠状面视图中 PRV 和 RV 逆行标记神经元(彩色点)的全脑图沿头尾方向总结(Fig. 2i)。此外,两种病毒在整个大脑中的半定量标记差异如 Fig. 2j 所示。Table 1 列出了大脑结构的缩写全名。总之,这些研究结果表明,PRV 和 RV 属于不同的病毒属并包含不同的病毒结构,当注射到腘淋巴结时,表现出不同的神经嗜性作用。
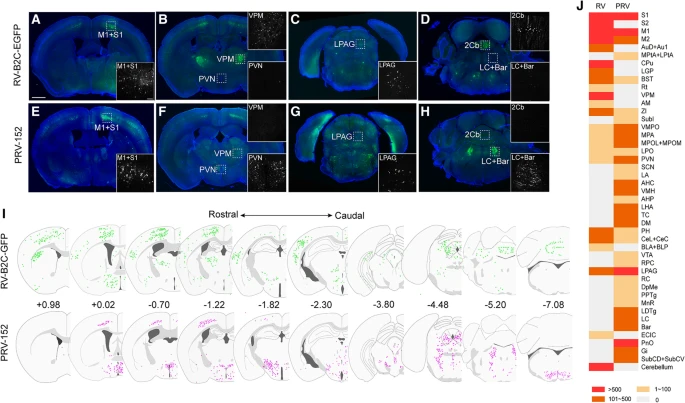
(a-d) RV-B2C-EGFP 在注射到淋巴结时逆行标记 M1 (A)、S1 (A)、VPM (B)、LPAG (C)、cerebellum (D),但不逆行标记 PVN、LC、Bar。
(e-h) PRV-152 在注射到淋巴结时逆行标记 M1 (E)、S1 (E)、PVN (F)、LPAG (G)、LC 和 Bar (H) ,但不逆行标记 VPM (F) 和 cerebellum(H)。
(i) 冠状视图中由 RV-B2C-EGFP(上行)和 PRV-152(下行)标记的 LHA 输入图沿头尾方向排列。逆行标记的神经元用彩色点表示。数字表示切片相对于 bregma 的位置(mm)。
(j) 全脑中 RV-B2C-EGFP 和 PRV-152 之间的半定量标记差异。
对每种病毒示踪剂分析了三只小鼠。插图显示了方框区域的放大图像。较低放大倍率图像中 Scale bars = 1 mm;较高放大倍率插图中 Scale bars = 100 um。
2. 不同单突触逆行示踪病毒的环路标记存在差异
PRV 和 RV 之间观察到的神经嗜性差异表明,由 PRV 或 RV 和其他常用替代方案设计的单突触逆行示踪剂(仅标记来自轴突末端靶向注射部位的直接输入神经元)将导致示踪环路差异。为了检验这一假设,我们比较了不同单突触逆行示踪剂识别的神经环路,包括源自 SAD-B19 疫苗株的糖蛋白缺失 RV (RV-ΔG)、TK(胸苷激酶)基因缺失 PRV (PRV-ΔTK-ΔUS9-EGFP)、逆转录腺相关病毒(rAAV2-retro)、retrobeads (Fig. 1b)。由于当 PRV-ΔTK-ΔUS9-EGFP 用于单突触逆行示踪时几乎检测不到荧光信号,可能是由于 TK 基因缺失导致复制缺陷,我们将其排除以进行进一步分析。
我们将 rAAV2-retro-YFP、RV-ΔG-EGFP 和 green retrobeads 注入下丘脑外侧区 (LHA)(Fig. 3a-a''' 中虚线圆圈区域),比较了三个示踪剂的 LHA 的直接上游环路。所有三种示踪剂均观察到内侧视前区 (MPA) 的逆行标记(Fig. 3b-b''' )。内侧前额皮质(mPFC)、无颗粒岛叶皮质(AI)(Fig. 3c-c' )、海马 CA1、基底外侧杏仁核(BLA+BLP+BLV)和基底内侧杏仁核(BMA+BMP)(Fig. 3i)中荧光标记的神经元通过 rAAV2-retro-YFP 示踪观察到,而使用 RV-ΔG-EGFP 在这些区域中检测到很少的阳性神经元(Fig. 3e-e', 3i)。相反,伏隔核(NAc)、室旁核(PVN)(Fig. 3e''-f' )、中央杏仁核(CeC+CeM)和终纹床核(BNST)(Fig. 3i)被 RV-ΔG-EGFP 标记,但几乎没有被 rAAV2-retro-YFP 标记(Fig. 3c''-d',3i)。当使用 green retrobeads 时,还观察到 NAc 和 PVN 的逆行标记(Fig. 3g-h' )。对每个示踪区域中标记的神经元的数量进行半定量,并在三个示踪剂之间进行比较(Fig. 3i )。脑结构缩写全名列于 Table 1。
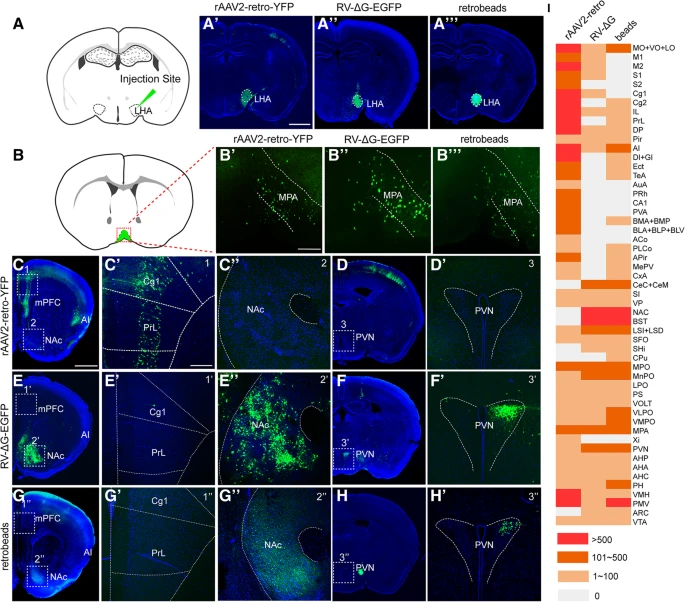
(a) 示踪剂注射到下丘脑外侧区(LHA)的示意图。
(a'-a''') rAAV2-retro-YFP、RV-ΔG-EGFP、green retrobeads 的注射位点被证实位于 LHA 中。
(b-b‴) 所有三种示踪剂均观察到 MPA 的逆行标记。
(c-d') 注射 rAAV2-retro-YFP 后,在 mPFC 中观察到逆行标记的神经元 (C, C'),而在 NAc (C, C'') 或 PVN (D, D') 中未观察到此类标记。
(e-f') 用 RV-ΔG-EGFP 逆行标记的神经元位于 NAc (E, E'') 和 PVN (F, F') 中,但在 mPFC (E, E') 中几乎观察不到。
(g-h′) green retrobeads 逆行标记 NAc (G, G'') 和 PVN (H, H'),但不逆向标记 mPFC (G, G')。
(i) 在三个示踪剂中比较 LHA 每个上游区域的标记神经元数量。
对每种病毒示踪剂或 retrobeads 分析了三只小鼠。对于 A′-A‴、C、D、E、F、G、H,Scale bars = 1 mm;放大图像 Scale bars = 200 μm。
由于 rAAV2-retro-YFP 感染的神经元中表达的 YFP 的亮度弱于 RV-ΔG-EGFP 感染的神经元中表达的 GFP(Fig. 3),因此存在 RV-ΔG-EGFP 阳性神经元但 rAAV2-retro-YFP 呈阴性可能是由于 LHA 上游区域 YFP 表达较低所致。为了排除这种可能性,我们将 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-Cre-tagBFP 病毒注射到 Ai9 报告小鼠的 LHA 中(Additional file 1: Figure S1A-S1C),其中即使是低 Cre 重组酶表达也可以通过 Cre/LoxP 系统在受感染的神经元中触发强而稳定的 tdTomato 表达。尽管相对于使用 rAAV2-retro-YFP 观察到的结果,rAAV2-retro-Cre-tagBFP 介导的 tdTomato 表达确实可以提高 rAAV2-retro-YFP 阴性大脑区域(包括 NAc 和 PVN)的标记效率(Fig. 3c-d', Additional file 1: Figure S1D-S1E'),这些区域中 tdTomato 阳性神经元的数量仍然远低于 RV-ΔG 标记的数量(Fig. 3e-f′, Additional file 1: Figure S1D-S1E', Additional file 1: Figure S1F-S1G')。此外,我们观察到 RV-ΔG-Cre-tagBFP 和 RV-ΔG-EGFP 示踪结果之间没有显着差异,可能是由于 RV 系统中基因表达水平较高(Fig. 3e-f' and Additional file 1: Figure S1F-S1G')。使用此 Cre/LoxP 系统,我们仍然观察到 RV-ΔG-Cre-tagBFP 和 rAAV2-retro-Cre-tagBFP 病毒示踪的环路之间存在显着差异(Additional file 1: Figure S1D-S1G'),这些差异与使用 rAAV2-retro-YFP 和 RV-ΔG-EGFP 观察到的标记模式是一致的。例如,当使用 rAAV2-retro-Cre-tagBFP 时,在 mPFC 中观察到强标记;当使用 RV-ΔG-Cre-tagBFP 时,在 NAc 和 PVN 中观察到强标记。为了进一步排除神经嗜性差异是由注射偏差差异引起的可能性,我们将 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 的混合物注射到 Ai9 报告小鼠的 LHA 中(Fig. 4a)。在此方案下,我们还获得了与分别注射两种病毒时一致的结果(Fig. 3, Additional file 1: Figure S1, and Fig. 4b-e'' )。
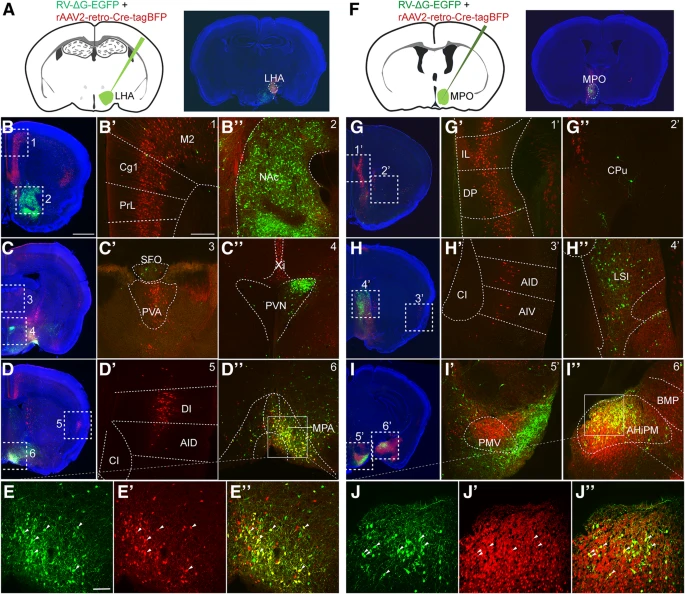
(a) Ai9 报告小鼠 LHA 中病毒共注射示意图及注射部位确认。
(b-e'') 当 LHA 同时注射 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 时,观察到两种病毒逆行标记的输入环路存在差异。mPFC (Cg1+PrL)、M2、PVA、DI 专门由 rAAV2-retro-Cre-tagBFP 标记,而 NAc 和 PVN 优先由 RV-ΔG-EGFP 标记,尽管在 MPA 中观察到重叠标记。
(f) 内侧视前核(MPO)中病毒共注射的示意图和注射位点。
(g-j'') 在 MPO 中注射 rAAV2-retro-Cre-tagBFP 与 mPFC、AI、PMV 中的优先标记相关。相反,在 MPO 中注射 RV-ΔG-EGFP 与 LSI 中的优先标记相关。在 AHiPM 中观察到重叠信号。
红色,由 rAAV2-retro-Cre-tagBFP 标记细胞表达的 tdTomato;绿色,由 RV-ΔG-EGFP 标记细胞表达的 GFP;蓝色,DAPI。A-D 和 F-I 的 Scale bars = 1 mm;B'-D', G'-I', B''-D'' and G''-I'' 的 Scale bars = 200 μm;E-E'' 和 J-J'' 的 Scale bars = 100 μm。
为了确定这种差异是否仅发生在 LHA 上游的环路内,我们还将这些示踪剂注入内侧视前核 (MPO)。同样,两种病毒之间的 MPO 标记环路也存在明显差异。如 Figure 4f-j'' 所示,mPFC 和前乳头核腹侧部分(PMV)仅被 rAAV2-retro-Cre-tagBFP 强烈标记,而侧隔核中间部分(LSI)主要被 RV-ΔG-EGFP 标记。值得注意的是,两种病毒在注射到 LHA 时都会标记内侧视前区,在注射到 MPO 时会标记杏仁核,其中标记的神经元彼此之间表现出很大的重叠(Fig. 4d-e'' and Fig. 4i-j'' )。为了根据上述结果分析不同逆行病毒示踪剂的示踪偏好,我们总结了整个大脑中由 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 标记的 LHA 和 MPO 的所有输入。大脑结构缩写全名列于 Table 1。如 Fig. 5a-c 所示,rAAV2-retro-Cre-tagBFP 优先标记大脑皮层,而 RV-ΔG-EGFP 优先标记基底神经节和下丘脑。在神经环路研究中,这种标记偏好将为正确使用 rAAV2-retro 和 RV-ΔG 示踪剂以达到最佳标记提供重要指导。值得注意的是,我们甚至观察到当 rAAV2-retro 和 RV-ΔG 被(共同)注射到背外侧膝状核(dLGN)(Fig. 5f)以标记初级视觉的皮质(V1)丘脑神经元时,它们对特定皮质层有显着的偏好。如 Figure 5d 所示,当使用 rAAV2-retro-YFP 时,阳性标记主要出现在 V1 的第五层(L5);而当使用 RV-ΔG-EGFP 和 green retrobeads 时,V1 中的阳性标记大部分位于第六层(L6)内。将 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 进一步共注射到 dLGN 中,证实了这两种病毒示踪剂的层偏好(Fig. 5e)。
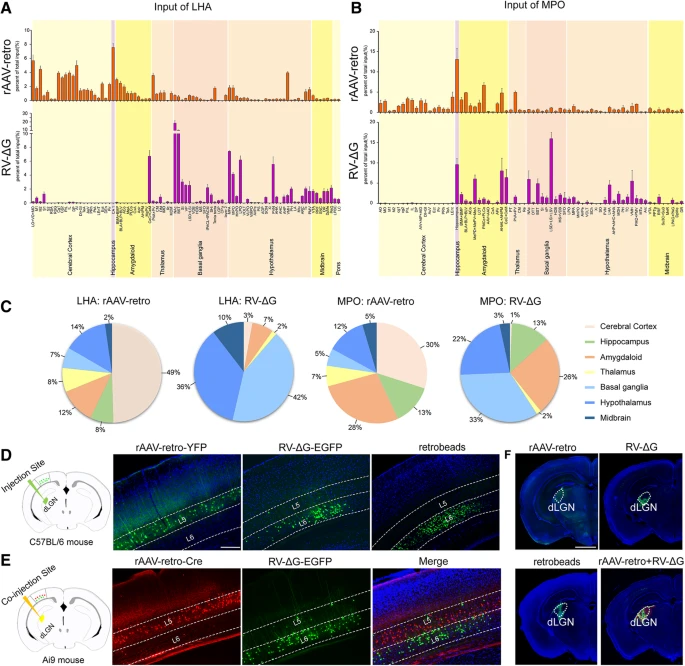
(a-b) 注射 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 后 LHA (A) 和 MPO (B) 输入环路逆行标记的全脑定量分析。对注射部位同侧的每个核中标记的神经元的数量进行量化,并将其归一化为每只小鼠(n = 3)的同侧标记神经元的总数。
(c) LHA 和 MPO 的标记输入脑区域的百分比以饼图表示,以表明大脑皮层中 rAAV2-retro 以及基底神经节和下丘脑中 RV-ΔG 的标记偏好。
(d) 将 rAAV2-retro-YFP、RV-ΔG-EGFP、green retrobeads 逆行示踪剂分别注射到 dLGN 中,用于初级视觉皮层(V1)中上游皮质丘脑神经元的逆行标记。rAAV2-retro-YFP 标记的 V1 皮质丘脑神经元主要位于第五层 (L5)。相比之下,RV-ΔG-EGFP 和 green retrobeads 主要标记 V1 第六层 (L6) 的皮质丘脑神经元。
(e) 将 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 的逆行示踪剂共同注射到 Ai9 小鼠的 dLGN 中,以同时标记 V1 中的上游皮质丘脑神经元。rAAV2-retro-Cre-tagBFP 主要标记 V1 第五层 (L5) 的皮质丘脑神经元,而 RV-ΔG-EGFP 主要标记 V1 第六层 (L6) 的皮质丘脑神经元。
(f) rAAV2-retro-YFP、RV-ΔG-EGFP、green retrobeads 的注射位点,以及 rAAV2-retro-Cre-tagBFP 与 RV-ΔG-EGFP 的共注射。
Scale bars = 200 μm in D-E, 1 mm in F。
考虑到不同逆行病毒示踪剂之间的标记差异,单独使用 rAAV2-retro 或 RV-ΔG 逆行示踪只能揭示部分直接上游区域。因此,有必要结合两个示踪剂的示踪数据以获得某个核的更全面的输入映射。对此,我们收集了 rAAV2-retro 和 RV-ΔG 的所有示踪数据,描绘了更全面的 LHA 直接上游环路图,为探索 LHA 环路的功能提供了重要参考,例如进食、唤醒、疼痛感知、体温调节、消化功能(Additional file 2: Figure S2)。
3. 通过 EnvA-TVA 伪型系统在 RV 抗性区域使用 RV-ΔG 进行逆行标记
上述结果表明,不同的逆行病毒与差异性环路标记有关,这很可能是由于病毒入侵的特异性受体的差异表达所致。LHA 上游 mPFC 区域中 RV-ΔG-EGFP 的稀少感染(Figs. 3, 4, 5, Additional file 1: Figure S1)可能是由于这条环路的 mPFC 神经元中缺乏 RV 受体。为了验证这一假设,我们采用了腺相关病毒 (rAAV9-EGFP-TVA) 表达的禽肉瘤白血病病毒受体 TVA 和禽肉瘤白血病病毒包膜蛋白 (EnvA) 假型 RV-ΔG (EnvA-RV-ΔG-DsRed) )以启用来自 LHA 的 mPFC 的 RV-ΔG 标记(Fig. 6)。为此,首先将 rAAV9-EGFP-TVA 注射到 mPFC 中,以在 mPFC 神经元及其末梢中过度表达 TVA。然后,EnvA-RV-ΔG-DsRed 被注入 LHA(Fig. 6h)。与来自 LHA 的 mPFC 中稀少的 RV-ΔG-DsRed 标记相比(Fig. 6a-g),mPFC 神经元在 LHA 的末端表达 TVA 确实可以导致成功标记 EnvA-RV-ΔG-DsRed 这些 mPFC RV 抵抗神经元(Fig. 6h-n)。根据这一发现,我们观察到 mPFC 中大量 DsRed 阳性神经元与 rAAV9-EGFP-TVA 感染的神经元重叠(Fig. 6i-l)。作为对照,我们进行了相同的实验,但没有将 TVA 注射到 mPFC 中,随后 LHA 中没有神经元被 EnvA-RV-ΔG-DsRed 标记(Fig. 6o-u)。同样,V1 中对 RV-ΔG 具有抗性的 L5 皮质丘脑神经元也可以通过 TVA 受体互补到 V1 皮层而被 dLGN 的 EnvA-RV-ΔG-DsRed 成功标记(Additional file 3: Figure S3)。综上所述,这些数据表明,TVA 受体的外源表达确实可以使 EnvA-pseudotyped RV 成功进入 RV 抗性神经元的轴突,支持了这些神经元对 RV-ΔG 的抗性是由于缺乏 RV 受体的观点。
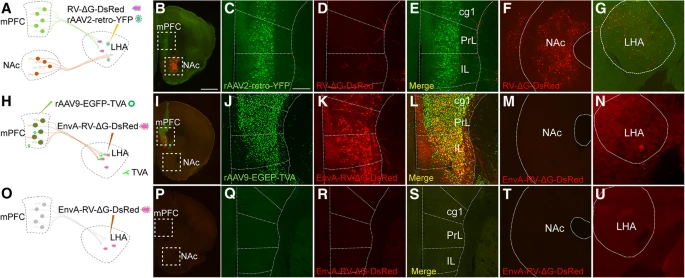
(a) 描绘 rAAV2-retro-YFP 和 RV-ΔG-DsRed 共注射到 LHA 中的示意图。
(b-f) 将 rAAV2-retro-YFP 和 RV-ΔG-DsRed 共注射到标记为不同输入核的 LHA 中。mPFC 由 rAAV2-retro-YFP 标记,NAc 由 RV-ΔG-DsRed 标记。
(g) LHA 中 rAAV2-retro-YFP 和 RV-ΔG-DsRed 的注射位点。
(h) 示意图描绘了注射表达 TVA 的辅助 AAV (rAAV9-EGFP-TVA) 以实现 mPFC 中的 EnvA pseudotyped RV (EnvA-RV-ΔG-DsRed) 感染。
(i-m) mPFC 神经元中 TVA 受体表达的互补使得 mPFC 中的 EnvA-RV-ΔG-DsRed 感染成为可能。 NAc 中未检测到 EnvA-RV-ΔG-DsRed 的阳性标记。
(n) LHA 中 EnvA-RV-ΔG-DsRed 的注射部位。
(o) 示意图描绘了在没有辅助 rAAV9-EGFP-TVA 的情况下将 EnvA-RV-ΔG-DsRed 注射到 LHA 中。
(p-t) 单独注射 EnvA-RV-ΔG-DsRed 后,在 mPFC 中未观察到信号。
(u) LHA 中 EnvA-RV-ΔG-DsRed 的注射部位。
B、I、P 中 Scale bars = 1 mm;放大图像中 Scale bars = 200 μm。
4. 通过单细胞转录组测序筛选有助于神经嗜性的候选基因
为了研究不同逆行病毒示踪剂标记的神经元的异质基因表达谱,这可能导致观察到的病毒神经嗜性差异,我们对受感染的神经元进行了单细胞转录组测序(Fig. 7a)。首先,将 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 分别平行注射到 Ai9 报告小鼠的下丘脑、丘脑、海马和杏仁核的不同核团中。然后将逆行标记的神经元进行单神经元分离,随后收集 29 组 rAAV2-retro-Cre-tagBFP 标记的神经元和 30 组 RV-ΔG-EGFP 标记的神经元。为了保证测序数据的可靠性,我们使用 25% 的数量阈值对样本进行过滤,并进一步分析了 21 组 rAAV2-retro-Cre-tagBFP 标记的神经元和 22 组 RV-ΔG-EGFP 标记的高质量神经元,其中每个 scRNA-seq 数据中捕获了超过 10,000 个基因转录本(Additional file 4: Figure S4A)。
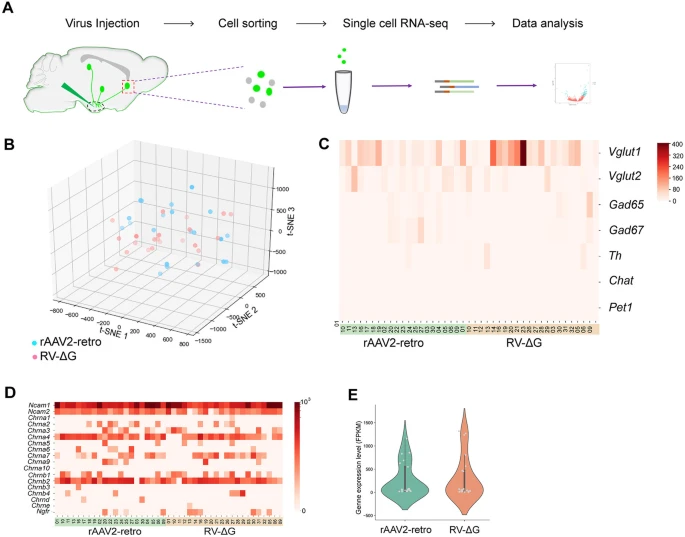
(a) 单细胞分离和 RNA 测序的流程图。
(b) rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 感染的神经元组的基因聚类分析。
(c) rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 组中神经元 marker genes 的表达,包括 VLGUT1、VGLUT2、GAD65、GAD67、TH、ChAT 和 Pet1。
(d) rAAV2-retro 和 RV-ΔG 标记组的每个样本中狂犬病病毒潜在受体候选者的表达热图,包括 Ncam、Chrna 和 Ngfr。
(e) rAAV2-retro 和 RV-ΔG 标记组之间 Ncam1 表达没有显着差异,P=0.088。
Chrna,烟碱乙酰胆碱受体基因;Ncam,神经细胞粘附分子基因;Ngfr:神经生长因子受体
经过基因测序和比对后,我们对 scRNA-seq 表达数据进行 t-SNE 降维分析,根据 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 感染的神经元的基因表达谱对它们进行聚类。如 Fig. 7b 所示,彩色点代表不同组的 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 感染的神经元,根据其基因谱进行分配,这表明 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 标记的神经元组之间没有明显的分布模式 。接下来,我们分析了不同类型神经元的特定 marker genes 的表达。rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 组之间没有观察到这些基因表达的显着差异(VLGUT1:P=0.5134,VGLUT2:P=0.5327,GAD65:P=0.1138,GAD67:P=0.1217, TH:P=0.2231,ChAT:P=0.3266,Pet1:P=0.3579),这表明 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 都可以感染所有类型的神经元(Fig. 7c)。
已报道狂犬病病毒的几种潜在候选受体,包括烟碱乙酰胆碱受体 (nAChR)、神经细胞粘附分子 (NCAMs) 和 p75NTR/Ngfr。然后,我们分析了所有 RV-ΔG-EGFP 和 rAAV2-retro-Cre-tagBFP 标记神经元组中 RV 受体候选基因的表达(Fig. 7d)。我们的数据表明,所有 RV 感染神经元组中仅 Ncam1 基因表达(Fig. 7d)。然而,RV-ΔG-EGFP 和 rAAV2-retro-Cre-tagBFP 标记神经元之间的 Ncam1 表达没有观察到显着差异(Fig. 7e)。同时,我们没有在 RV-ΔG 感染的神经元组中观察到 Ncam、nAChR 和 Ngfr 基因的排他性表达(Fig. 7d),这意味着这些候选受体可能不是 RV-ΔG 神经嗜性的重要因素。
由于介导病毒入侵的受体理论上应该是存在于细胞膜上的突触特异性蛋白,因此潜在的病毒受体候选基因很可能满足以下三个标准:(1)在特定病毒标记的所有神经元组中表达; (2) 具有膜特异性; (3) 具有突触特异性。因此,我们开发了一个生物信息学分析流程来筛选潜在的病毒受体,如 Additional file 4: Figure S4B 所示。最后,我们分别鉴定了在所有 rAAV2-retro-Cre-tagBFP 和 RV-ΔG-EGFP 组中表达的 682 和 527 个突触特异性膜蛋白基因(Additional file 4: Figure S4C)。基因功能(GO)分析显示,受体候选基因在多个通路中富集,包括"细胞投射"、"突触间隙"、"细胞-细胞连接"、"突触后" 通路(Additional file 4: Figure S4D-S4E)。
5. rAAV2-retro 和 RV-ΔG 示踪剂的神经毒性差异
由于嗜神经病毒也被广泛应用于环路功能的记录以及环路操作和基因谱研究,我们的目的是系统地分析这些病毒在注射部位和逆行标记细胞核中的神经毒性。首先,注射 2 周后从实验组和对照组中解剖注射部位(LHA)、rAAV2-retro-YFP 标记的细胞核(mPFC)、RV-ΔG-EGFP 标记的细胞核(NAc)(Fig. 8a)。然后从不同细胞核的组织中提取 RNA 并进行文库构建和 RNA 测序。对不同群体基因谱的生物信息分析表明,逆行病毒示踪剂的感染确实可以导致基因表达的改变,如 Fig. 8b-e 中的火山图所示。与 rAAV2-retro-YFP 感染相比,RV-ΔG-EGFP 感染在注射部位和逆行标记细胞核中引起更大的基因表达变化,表明 RV-ΔG-EGFP 感染对基因表达的影响更明显(Fig. 8b-e)。我们还对所有组中差异表达的基因进行了基因功能(GO)分析,结果表明 RV-ΔG-EGFP 感染主要影响与免疫反应、细胞粘附、抗原加工/呈递相关的基因,在注射部位以及逆行标记的细胞核(Fig. 8f-g)。相反,在 rAAV2-retro 注射后,在注射位点或逆行标记的细胞核中没有富集与特定术语相关的基因。因此,我们随后对各组差异表达基因进行了 KEGG 通路分析,其数据显示在 Additional file 5: Figure S5 中。
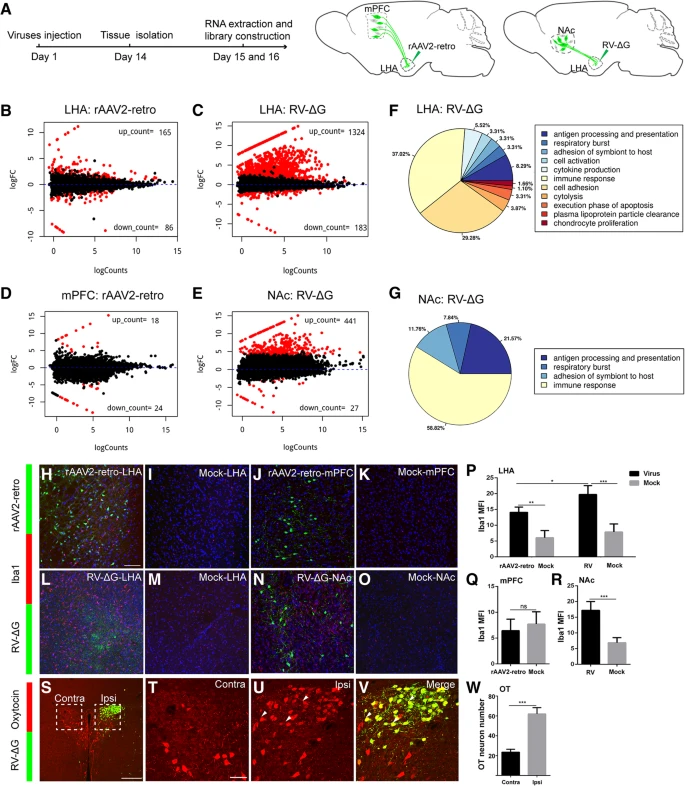
(a) 病毒注射和组织提取用于 RNA 测序的示意图。
(b) 将 rAAV2-retro-YFP 注射到 LHA 中会导致基因表达谱发生变化。
(c) 将 RV-ΔG-EGFP 注射到 LHA 中会导致基因表达谱发生变化。
(d) 将 rAAV2-retro-YFP 注射到 LHA 后,逆行标记的 mPFC 中的基因谱发生变化。红点代表有显着变化的基因,黑点代表没有显着变化的基因。
(e) 将 RV-ΔG-EGFP 注射到 LHA 后,逆行标记的 NAc 中的基因谱发生变化。
(f-g) RV-ΔG-EGFP 注射位点 (LHA) 和逆行标记核 (NAc) 差异表达基因的 GO-term 通路分析。
(h-i) rAAV2-retro-YFP 组中小胶质细胞标记物 Iba1 的免疫染色。在注射部位观察到 Iba1 阳性细胞,但在注射 PBS(模拟)的 LHA 中几乎没有观察到。
(j-k) 将 rAAV2-retro-YFP 或 PBS(模拟)注射到 LHA 后,在同侧 mPFC 中未检测到激活的小胶质细胞。
(l-m) 当 RV-ΔG-EGFP 注射到 LHA 中时,在注射部位观察到强烈的小胶质细胞激活,而在注射 PBS 的(模拟)LHA 中观察到很少的信号。
(n-o) 将 RV-ΔG-EGFP 注射到 LHA 后,在逆行标记的同侧 NAc 中观察到强烈的小胶质细胞激活,但在模拟对照的同侧 NAc 中没有观察到。
(p) 病毒注射后 LHA 中 Iba1 的平均荧光强度 (Mean±SEM) 的定量。rAAV2-retro-YFP: 14.09±0.8373, Mock: 6.059±1.132, rAAV2-retro-YFP vs Mock: P=0.0013; RV-∆G-EGFP: 19.75±1.403, Mock: 7.885±1.265, RV-∆G-EGFP vs Mock: P = 0.0008; rAAV2-retro-YFP vs RV-∆G-EGFP: P = 0.0133;n = 4,n 为小鼠数量。
(q) 注射 rAAV2-retro-YFP 和 PBS 的 mPFC 中 Iba1 的平均荧光强度 (Mean±SEM) 的定量。 rAAV2-retro-YFP: 6.438±1.114, PBS: 7.713±1.192, P = 0.4639, n = 4。
(r) 注射 RV-ΔG-EGFP 和 PBS 的 NAc 中 Iba1 的平均荧光强度 (Mean±SEM) 的定量。RV-∆G-EGFP: 17.20±1.408, Mock: 6.883±0.8132, P = 0.0007; n = 4。
(s) 将 RV-ΔG-EGFP 注射到 LHA 后 7 天,对 PVN 神经元中的催产素(oxytocin)进行免疫染色。 (t, u) S 中的方框区域被放大,以显示病毒注射侧的同侧和对侧 PVN 中的催产素阳性神经元。
(v) 虚线框的高倍放大图像,用绿色和合并颜色描绘 S 中的同侧 PVN。绿色,用 RV-ΔG-EGFP 表达的 GFP;红色,催产素免疫染色信号。
(w) 同侧和对侧 PVN 中催产素阳性神经元的定量 (Mean±SEM)。Contra: 23.50±1.443, Ipsi: 62.00±3.136, P=0.0001, n = 4。
MFI,平均荧光强度;Ipsi,同侧;Contra,对侧。n 是小鼠数量。S 的 Scale bars =200 μm;H-O 的 Scale bars = 100 μm;T-V 的 Scale bars = 50 μm。Unpaired t-tests, *P < 0.05, *
由于中枢神经系统中的病毒感染可以激活小胶质细胞,我们进行了 Iba1 免疫组织化学来评估注射部位和逆行标记细胞核中 RV-ΔG 和 rAAV2-retro 感染诱导的小胶质细胞激活。如 Figure 8l-o, p, r, Additional file 6: S6C-S6D 所示,RV-ΔG 注射后,注射部位 (LHA) 和逆行示踪区域 (NAc) 的小胶质细胞浸润显着增加。同时 rAAV2-retro 感染引起的小胶质细胞激活明显较轻(Fig. 8h-k, p, q, Additional file 6: Figure S6A-S6B),即使是在 30 天后,这表明 RV-ΔG 对注射部位和逆行标记区域的毒性更大。相反,在注射部位和逆行标记的细胞核中,RV-ΔG 和 rAAV2-retro 感染中,星形胶质细胞标记物 GFAP(胶质纤维酸性蛋白)没有观察到显着变化(Additional file 6: Figure S6E-S6O)。我们通过对不同激素的基因表达进行免疫染色分析,进一步研究了 RV 感染对 PVN(分泌各种神经内分泌激素的中央核)神经元的影响。因此,我们分别在 LHA 中注射 RV-ΔG-EGFP 后 7 天和 14 天检查了 PVN 中催产素和加压素的表达。我们的研究结果表明,超过 90% 的 RV 标记的 PVN 神经元表达催产素,并且 RV 注射部位同侧的 PVN 区域,催产素阳性神经元的数量显着增加,相对于对侧观察到的数量 (Fig. 8s-w, Additional file 6: Figure S6P-S6T)。这些发现表明 RV 感染可导致 PVN 神经元异位催产素表达。相反,RV-ΔG-EGFP 感染不会改变加压素的表达(Additional file 6: Figure S6U-S6Y)。
6. 通过组合不同的病毒示踪剂进行针对特定目标的高阶环路示踪的概念验证
我们的数据表明,应用 rAAV2-retro 或 RV-ΔG 进行逆行示踪仅揭示了直接上游区域的一部分。因此,为了获得涉及特定靶核的高阶环路的更全面的图谱,我们提出了一种结合 rAAV2-retro、rAAV 和 rRV 示踪剂的策略,如 Fig. 9a 所示。在该策略中,rAAV2-retro-Cre-tagBFP 和 RV-ΔG-Cre-tagBFP 首先分别注射到目标核 (i) 中,以示踪并将 Cre 递送到直接上游连接的核 (ii)。Cre 依赖性 rAAV9-DIO-EGFP-TVA 和 rAAV9-DIO-RVG 以及 EnvA-RV-ΔG-DsRed 被注射到 rAAV2-retro-Cre-tagBFP 标记的核 ii 中,用于二次输入的跨单突触标记环路。在 RV-ΔG-Cre-tagBFP 标记的上游核 ii 中,rAAV9-DIO-RVG 被注入核 ii 中,以确保 RV 能够跨突触跳跃到目标 I 的二阶环路。因此,我们将 rAAV2-retro-Cre-tagBFP 注射到 LHA 中,并将 rAAV9-DIO-EGFP-TVA 和 rAAV9-DIO-RVG 的组合注射到 mPFC 中(Fig.9b)。14 天后,EnvA-RV-ΔG-DsRed 被注入 mPFC 以追踪次级上游环路(Fig.9b)。如 Figure 9c-d 所示,rAAV2-retro-Cre-tagBFP 特异性标记针对 LHA 的 mPFC 神经元,并触发这些神经元中 TVA 和 RVG 的表达,从而介导 EnvA-RV-ΔG-DsRed 的感染并进一步启动 LHA 次级上游环路的跨单突触示踪。在局部 mPFC、无颗粒岛叶皮层 (AI)、外侧眶皮层 (LO)、梨状皮层 (Pir)、前内侧丘脑核 (AM) 和基底外侧杏仁核(BLA)观察到被 EnvA-RV-ΔG-DsRed 感染的 DsRed 阳性神经元,为 LHA 提供二阶输入(Fig. 9c-g)。
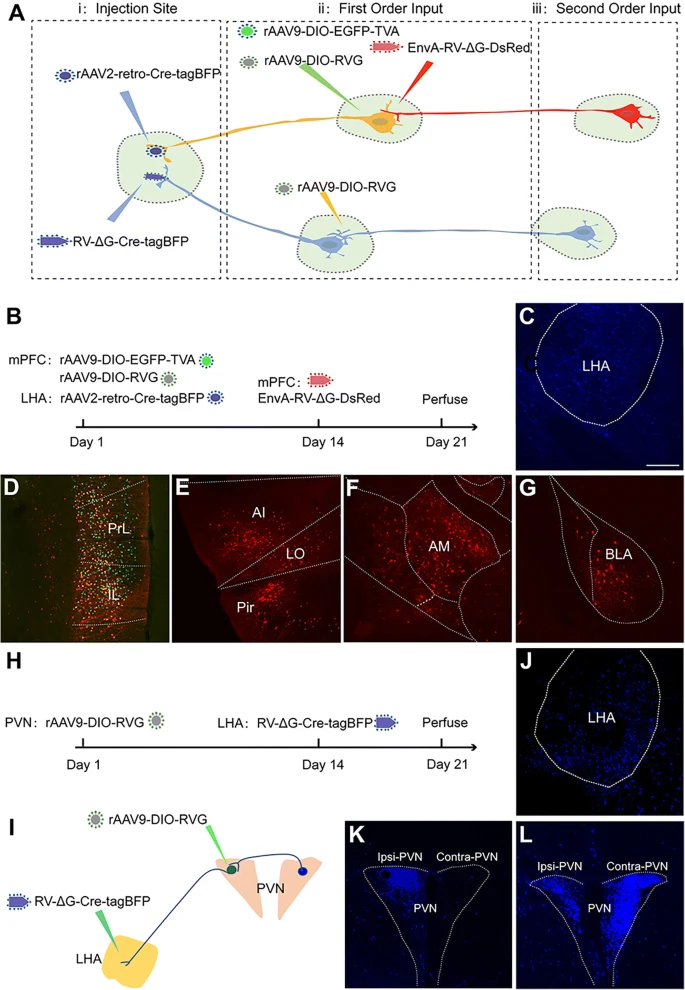
(a) 通过组合不同的病毒示踪剂来绘制目标核的高阶输入环路的策略。
(b) 结合 rAAV2-retro-Cre-tagBFP、rAAV9-DIO-EGFP-TVA、rAAV9-DIO-RVG 和 EnvA pseudotyped RV-ΔG-DsRed 的 LHA 高阶环路绘制流程图。
(c) LHA 中 rAAV2-retro-Cre-tagBFP 的注射位点。
(d) 来自 LHA(一级)的直接上游神经元在 mPFC 的 PrL 和 IL 中用绿色和红色荧光标记(也称为起始神经元)。绿色,TVA 阳性神经元;红色,EnvA-RV-ΔG-DsRed 阳性神经元;黄色,起始神经元。
(e-g) 起始神经元上游的细胞核(二阶)在 AI、LO、Pir (E)、AM (F) 和 BLA (G) 中仅用红色荧光标记。
(h-i) 通过在 PVN 中组合 rAAV9-DIO-RVG 和 RV-ΔG-Cre-tagBFP 标记获得 LHA 高阶环路的流程图。
(j) RV-ΔG-Cre-tagBFP 的注射位点。
(k) RV-ΔG-Cre-tagBFP 仅逆行标记同侧 PVN 中的神经元。
(l) 通过将 AAV-DIO-RVG 注射到同侧 PVN 中来补充 RV-G,导致对侧 PVN 中的二级神经元被标记。Scale bars = 200 μm。
此外,为了绘制由 RV-ΔG 示踪的 LHA 的高阶上游环路,我们在将 RV-ΔG-Cre-tagBFP 注射到 LHA 之前 14 天将 rAAV9-DIO-RVG 注射到 PVN 中(Fig. 9h-j)。如 Figure 9k-l 所示,靶向 LHA 的 PVN 神经元被 RV-ΔG-Cre-tagBFP 逆行标记,导致这些神经元中 RV-G 的表达,并启动对 PVN-LHA 环路内上游核的跨单突触示踪,在 RV-ΔG-Cre-tagBFP 注射后 7 天后(例如,对侧 PVN)。总而言之,这些数据为通过 rAAV2-retro、rAAV 和 rRV 的组合对特定目标核进行更全面的高阶环路绘制提供了概念验证。
讨论
病毒已经进化为通过不同的策略入侵中枢神经系统(CNS)。随着不同嗜神经病毒的各种潜在受体被发现,不同的病毒可能在神经系统中表现出不同的嗜性模式。在这方面,我们的研究结果表明,中枢神经系统内的逆行跨多突触示踪剂(例如伪狂犬病病毒(PRV)和狂犬病病毒(RV))的嗜性确实存在差异。因此,从这些嗜神经病毒修饰的单突触示踪剂原则上应该表现出对特定环路的优先标记。在这项研究中,我们设计了表达 GFP 的 TK 缺失 PRV (PRV-ΔTK-ΔUS9-EGFP) 和糖蛋白缺失 (ΔG) RV,并比较了它们在啮齿动物大脑中的神经嗜性作用。然而,PRV-ΔTK-ΔUS9-EGFP 的荧光信号几乎检测不到,很可能是由于 TK 缺失后病毒复制受损。因此,我们关注最广泛使用的逆行单突触示踪剂:糖蛋白缺失 (ΔG) RV、rAAV2-retro 和 retrobeads。我们的数据显示这些示踪剂确实表现出优先逆行环路标记。投射到外侧下丘脑区(LHA)和内侧视前核(MPO)的 rAAV2-retro 标记神经元优先位于大脑皮层。相反,那些被 RV-ΔG 标记的主要位于基底神经节和下丘脑。
值得注意的是,我们证明 rAAV2-retro 主要标记来自背外侧膝状核 (dLGN) 的初级视觉皮层 (V1) 的第 5 层 (L5),而 RV-ΔG 和 green retrobeads 主要标记 V1 的 L6。由于当这两种病毒共同注射时也观察到这种偏好,因此不同的标记模式并不是由于不同病毒的注射变化造成的。RV-ΔG 和 retrobeads 对 V1 第 6 层皮质丘脑神经元的标记与之前的研究结果一致,其中类似的 RV-ΔG 或 retrobeads 示踪剂被注射到 dLGN 中并专门标记 V1 的第 6 层皮质丘脑神经元。很少有研究报告从 dLGN 标记 V1 中的第 5 层皮质丘脑神经元,这可能是由于当前环路示踪剂的限制,但已经证明 V1 皮质的第 5 层神经元可以投射到 dLGN。同时,V1 中添加 TVA 有助于注射到 dLGN 中的 EnvA pseudotyped RV-ΔG 示踪第 5 层皮质丘脑神经元,进一步证实了 V1 第 5 层神经元对 dLGN 的投射,也支持了神经元对 RV-ΔG 抵抗的观点是由于缺乏 RV 受体。
在单突触病毒示踪剂之间的神经性比较中,使用了具有不同启动子的 rAAV2-retro 病毒,包括 rAAV2-retro-YFP(ef1α 启动子)和 rAAV2-retro-Cre-tagBFP(CMV 启动子)。尽管两种 rAAV2-retro 病毒给出的结果基本一致,但仍然存在轻微差异,例如 rAAV2-retro-CMV-Cre-tagBFP 而不是 rAAV2-retro-YFP 在 PVN、Xi nucleus、NAc 中进行标记。虽然不同的启动子(CMV 或 ef1α)可以在一定程度上影响基因表达,但这种标记差异最可能的原因可能是由于 Cre 引起的扩增效应。然而,不同的启动子并不影响 rAAV2-retro 示踪剂的向神经嗜性及其与 RV-ΔG 示踪剂相比的标记差异。
这些示踪剂在中枢神经系统中的独特嗜性可能是由于启动它们进入神经元的相应受体的存在差异所致。据报道,犬腺病毒 2 型(CAV2)通过与柯萨奇病毒和腺病毒受体(CAR)结合来感染细胞。CAR 受体的互补可以提高 CAV2 逆行标记效率。同时我们的数据表明,TVA 受体的外源表达确实可以使 EnvA-pseudotyped RV 成功进入 RV 抵抗神经元的轴突。虽然包括 nAChR、NCAM 和 p75NTR/Ngfr 在内的几个基因被认为是潜在的 RV 受体,但我们没有观察到 rAAV2-retro 和 RV-ΔG 组之间这些基因的表达水平存在显着差异。这些 RV 感染的潜在受体可能不是中枢神经系统中 RV 神经嗜性的决定性因素。根据嗜性差异可能是由于病毒受体表达差异造成的这一观点,我们发现,RV-ΔG 抗性核中 TVA 受体的外源表达确实可以使该核中的 EnvA-pseudotyped RV 感染。为了研究病毒嗜性的分子机制,我们利用高质量的单细胞 RNA 序列数据分析了 22 组 RV-ΔG 感染的神经元和 21 组 rAAV2 逆转录病毒感染的神经元的基因表达谱。理论上,潜在的病毒受体应该是在所有标记神经元中表达的突触膜蛋白。因此,我们使用不同的数据库筛选了 RV 和 rAAV2-retro 候选受体基因,并鉴定了 527 个基因作为 RV 入侵的潜在受体和 682 个基因作为 rAAV2-retro 感染的潜在受体。此前的研究表明,病毒受体以外的因素也可能影响不同病毒的嗜性。Albisetti et al. 证明某些受体介导病毒进入背根神经节(DRG)的感觉神经元,并且宿主限制因素可能抑制病毒转录或复制,从而影响环路绘制的最终效率。因此,未来的研究应该采用其他生化和病毒学实验来鉴定这些病毒的确切受体或宿主限制因素,这可能有助于阐明病毒入侵的机制并改进基于病毒的逆行标记技术。
除了在神经环路绘制中的作用外,嗜神经病毒还与 Ca2+ 指示剂和光遗传学/化学遗传学技术相结合,广泛应用于特定神经环路的功能分析和操作。此外,表达荧光蛋白、标记核糖体亚基或其他功能蛋白的病毒示踪剂能够分离目标神经元,并对特定环路内的转录组、蛋白组和表观基因组进行单细胞测序分析。然而,不同的病毒在感染的不同阶段与宿主细胞中的神经毒性水平不同。因此,需要进一步的研究来更充分地降低不同病毒的毒性,以避免神经元操作、记录和基因测序过程中的误差。先前的研究表明,RV-ΔG 注射后 5-11 天神经元特征基本保持不变,因此可应用于此窗口期间的神经元活动记录和光遗传学操作。然而,注射 RV-ΔG 后 16 天,神经元开始死亡。在本研究中,我们使用更灵敏的方法:基因谱分析,研究了注射部位和逆行标记神经元的神经毒性。我们的数据表明,RV-ΔG 和 rAAV2-retro 对注射部位和逆行标记部位均产生一定影响,尽管这些变化对于 RV-ΔG 感染更为深远。因此,在功能记录、操作和基因图谱分析过程中应考虑此类影响。
在我们的研究中,RV-ΔG 和 rAAV2-retro 之间的神经嗜性和神经毒性的平行比较是在注射后 14 天进行的,此时病毒标记信号可以达到足够高的水平以进行检测。根据之前的出版物和我们的经验,RV-ΔG 标记的神经环路将在注射后 14 天左右达到优化结果,此时标记效率较高且毒性相对较低。对于 rAAV 和 rAAV-retro,注射后约 2-3 周即可达到有效的标记结果。因此,我们比较了病毒注射后两周时间窗口内的毒性,为使用具有可比交的示踪效果的不同环路示踪剂进行神经环路追踪引起的毒性提供一些参考。对于 PRV-ΔTK-ΔUS9-EGFP,我们推测 EGFP 无法检测到的表达主要是由于没有 TK 基因的复制缺陷导致 PRV 基因组的低拷贝,最近一篇论文显示没有 Cre 重组酶的 PRV-DIO-ΔTK-GFP 感染的输入神经元中几乎没有 GFP 荧光。
尽管 rAAV2-retro 具有较低的毒性水平,但它不能替代 RV-ΔG 进行环路追踪,因为与 RV-ΔG 相比,rAAV2-retro 表现出不同的嗜性模式,并且不能用于跨突触环路示踪。一些研究小组已经通过生成新的 RV 示踪剂成功降低了 RV 细胞毒性并增加了嗜性。CVS-N2cΔG RV 菌株、新设计的自失活 ΔG-RV (SiR) 和不含聚合酶基因的重组 RV 表现出神经元毒性的显着降低。源自固定狂犬病病毒株的新型缺失突变病毒示踪剂 CVS-N2cΔG 与疫苗株 SAD-ΔG 相比,对神经元的毒性较小。通过条件调节病毒核蛋白 (N) 稳定性来设计自失活 ΔG-RV (SiR) 示踪剂,或通过同时删除糖蛋白 (G) 和 RNA 聚合酶 (L) 来消除 RV 病毒基因组的复制基因,两者都显着降低毒性,更有利于神经环路功能操纵。然而,这些示踪剂的拯救效率低于疫苗株 SAD-ΔG,且这些病毒示踪剂的制备需要较高的专业知识和技术要求,限制了其在神经环路研究中的广泛应用。未来的研究需要更全面地阐明病毒复制和毒性的机制,然后通过病毒反向基因工程(例如精确控制细胞毒性相关基因的表达)来降低病毒示踪剂的毒性。
由于本研究中检查的不同病毒示踪剂表现出不同的嗜性模式,我们通过结合两种病毒示踪剂的逆行示踪数据,重建了更全面的 LHA 直接输入环路图。将该策略与光遗传学和电生理学等其他技术相结合,可能会极大地促进更全面地阐明细胞核的功能。此外,我们进一步提供了概念证明,即这些病毒的组合可用于描绘特定目标核的高阶输入。利用这种策略,研究人员可以创建更全面的直接上游环路以及相应的二阶环路图。最近病毒学、神经科学和生物技术(例如蛋白质工程和新功能指标的开发)领域的多学科工作表明,具有低毒性和多样化功能应用的可用神经示踪剂的数量在不久的将来将会增加。这些进步不仅将提高我们系统地绘制、记录和操纵神经环路活动的能力,而且还可能使我们能够阐明最复杂的大脑网络中环路的基因谱和(表观)遗传结构。
总结
在我们的研究中,我们系统地评估了不同跨多突触和单突触逆行病毒示踪剂(包括 PRV、RV 和 rAAV2-retro)的神经嗜性差异。单突触病毒示踪剂 rAAV2-retro 在大脑皮层中表现出更多的偏好,而 RV-ΔG 在基底节和下丘脑中表现出更多的偏好。这种不同的标记偏好表明,RV-ΔG 可以首先考虑映射来自基底神经节和下丘脑的输入,而 rAAV2-retro 可以考虑用于来自大脑皮层的输入标记。结合 RV-ΔG 和 rAAV2-retro 的数据将为感兴趣的脑核的精确神经环路描绘提供更全面的输入图。值得注意的是,当将它们应用于追踪 V1 皮质丘脑神经元至 dLGN 时,在第 5 层中的 rAAV2-retro 和第 6 层中的 RV-ΔG 观察到了皮质层偏好。RV-ΔG 和 rAAV2-retro 对 V1 皮质丘脑神经元的层特异性标记将用于剖析特定层中皮质丘脑神经元的环路图和功能。RV-ΔG 和 rAAV2-retro 均在注射部位和逆行标记部位产生神经毒性影响,RV-ΔG 感染的影响更为深远。特别是对于神经环路的功能研究,有必要考虑示踪病毒引起的神经毒性。最后,通过结合 rAAV2-retro、RV 和 rAAV 示踪剂,完成了对特定目标核进行更全面的高阶环路示踪的概念验证策略。我们的研究结果为病毒示踪剂在神经环路研究中的适当应用提供了重要参考。
注:本文为个人学习笔记,仅供大家参考学习,不得用于任何商业目的。如有侵权,请联系作者删除。

本文由 mdnice 多平台发布