文章信息
题目:Mitogen-activated protein kinase TaMPK3 suppresses ABA response by destabilising TaPYL4 receptor in wheat
刊名:New Phytologist
作者:Ying Liu,You-Zhi Ma, Zhao-Shi Xu et al.
单位:Institute of Crop Science
日期:20 June 2022
01
摘要
脱落酸 (ABA) 受体被认为是植物中 ABA 敏感性和水分生产力的靶向操纵。对其稳定性或活性的调节将直接影响 ABA 信号传导。丝裂原活化蛋白激酶 (MAPK) 级联连接多个环境和植物发育线索。然而,ABA 信号和 MAPK 级联相互作用的分子机制在很大程度上仍然难以捉摸。
TaMPK3过表达降低了干旱耐受性和小麦对 ABA 的敏感性,显着削弱了 ABA 对生长的抑制作用。在干旱胁迫下,过表达系的成活率、芽鲜重和脯氨酸含量较低,但在幼苗期丙二醛水平较高,并且在成年期温室和田间条件下的粒宽和千粒重均降低。TaMPK3 -RNAi 增加耐旱性。
TaMPK3 与 TaPYL4 的相互作用通过促进其泛素介导的降解导致 TaPYL4 水平降低,而 ABA 处理减少了 TaMPK3-TaPYL 相互作用。此外,在 ABA 处理下,过表达 TaMPK3 的小麦植物中 ABA 信号蛋白的表达受损。发现 MPK3-PYL 交互模块在单子叶植物和双子叶植物中是保守的。
我们的研究结果表明,MPK3-PYL 模块可以作为一种负调控机制,在小麦中平衡适当的干旱胁迫反应和正常的植物生长信号。
2
技术路线
小麦品种“Fielder”(野生型,WT)用于扩增基因序列(TaMPK3和TaPYLs)
产生TaMPK3转基因小麦植物
ABA敏感性测定
小麦叶肉原生质体中的亚细胞定位和双分子荧光互补 (BiFC) 测定
酵母二杂交 (Y2H) 测定
烟草中的荧光素酶互补成像 (LCI) 和 BiFC 检测
体外pull down试验
Co-IP 检测
体外磷酸化测定
干旱处理
03
主要结果
3.1 TaMPK3 与 TaPYL4 的互作
TaPYL4 先前已被证明参与调节小麦的耐旱性。为了拓宽我们对小麦中 PYL 的了解,TaPYL4 被用作 Y2H 筛选小麦 cDNA 文库的诱饵,该文库使用从干旱处理的小麦品种 Jimai60 的叶子中提取的 RNA 构建,以识别潜在的蛋白质相互作用。TaPYL4 的相互作用候选者列于表S6. 候选相互作用子的序列分析表明,MAPK 的 TaMPK3 表现出与 TaPYL4 的潜在直接相互作用。TaMPK3 包含典型的 MPK 特征基序,包括 ATP 结合特征 (IGRGAYGIVCSVMNFETREMVAIKK)、催化 C 环 (VIHRDLKPSNLLL)、激活 T 环 (TEY)、共同对接 (CD) 域 (LHDVADEPIC) 和 EF 手 CBP (RMLTFNPLQRITVEEAL)域(图S1)。与公共数据库 (NCBI) 的氨基酸序列比对显示,它编码的蛋白质与拟南芥 AtMPK3 (73.42%) 和水稻 OsMPK3 (91.33%) 具有高度序列相似性(表S7)。使用来自其他植物物种的 TaMPK3 及其直系同源物的全长氨基酸序列重建的系统发育树显示出来自单子叶植物和双子叶植物的 MPK3 之间的单独聚类,表明这些谱系之间存在潜在的功能分歧(图S2)。
进一步的证据表明,共表达 TaPYL4-AD 和 TaMPK3-BD 的酵母细胞能够在缺乏 Leu、Trp、Ade 和 His 的合成脱落选择培养基上生长(图 1a)。使用烟叶中的 LCI 分析,在 TaMPK3-nLUC/TaPYL4-cLUC 共注射区域可以观察到强发光信号,而在阴性对照的共注射区域未检测到信号(图1b)。然后,我们纯化了重组 GST 标记的 PYL4(TaPYL4-GST)和 MBP 标记的 TaMPK3(TaMPK3-MBP)用于 GST 下拉测定,这表明 TaPYL4-GST 蛋白可以下拉 TaMPK3-MBP 蛋白(图1c)。使用瞬时表达 TaPYL4-MYC 与 Flag 或 TaMPK3-Flag 的烟叶进行的进一步免疫共沉淀(Co-IP)测定证实 TaPYL4-MYC 可以被 TaMPK3-Flag 融合蛋白共免疫沉淀,但不能被 Flag 对照(图1d)。这些结果表明,TaPYL4 确实在体外和体内与 TaMPK3 发生物理相互作用。

编辑切换为居中
添加图片注释,不超过 140 字(可选)
Fig. 1
3.2 TaMPK3在体外直接与多个 TaPYL 相互作用
先前在小麦中至少报告了 9 种 TaPYL。鉴于这九种蛋白质之间的高度序列相似性(图S3)和相似的亚细胞位置(图S4),因此可能具有冗余功能,我们在小麦叶肉原生质体中进行了BiFC测定以确定TaPYL1-9是否可以物理相互作用TaMPK3。这些测定在与 TaMPK3-nYFP 和 TaPYLs-cYFP 共转化的小麦叶肉原生质体中显示出明显的 YFP 荧光,但在阴性对照中则没有(图 2a)。
为了验证这些结果,我们随后进行了 LCI、Y2H 和pull down分析。与 BiFC 实验一致,LCI 分析在共表达 TaMPK3-nLUC/TaPYLs-cLUC 的样品中显示出强烈的发光信号,但在阴性对照中没有,这表明 TaMPK3 可以与 TaPYL1-9 相互作用(图2b)。Y2H 测定表明,共表达 TaMPK3-BD 和 TaPYL4/5/6/7/8/9-AD 而不是 TaPYL1/2/3-AD 的酵母细胞可以在缺乏 Leu 的合成脱落选择培养基上生长, Trp、His 和 Ade(图2c),进一步表明 TaMPK3 与酵母中一组有限的 PYL 之间的相互作用。然后,我们用纯化的重组 GST 标记的 TaMPK3 (TaMPK3-GST) 和 His 标记的 TaPYL1-9 (TaPYLs-His) 进行了下拉分析,发现所有九种 TaPYL-His 蛋白但不是 His 标签可以拉下 TaMPK3-GST ,表明TaMPK3在体外与TaPYLs发生物理相互作用(图2d)。总之,小麦中报道的所有PYL蛋白(TaPYL1-9)都可以与TaMPK3相互作用,这表明TaMPK3可能具有控制小麦中TaPYL受体活性的保守调节功能。

编辑切换为居中
添加图片注释,不超过 140 字(可选)
Fig. 2
3.3 ABA 敏感性在 TaMPK3 过表达的小麦植物中降低,而在TaMPK3 - RNAi 小麦植物中增强
鉴于通过 Y2H 筛选获得的TaMPK3序列位于小麦的亚基因组 D,因此我们选择TaMPK3 -4D ( TraesCS4D02G198600 )由玉米 ( Z. mays ) 泛素启动子驱动。使用 qPCR 相对表达测定的定量证实TaMPK3 -OE 系(OE-2、OE-6 和 OE-11)表现出明显更高的转录水平(图S6a),而 RNAi 系(i-1、i-3、和 i-10)具有较低的TaMPK3表达(图S6b)。通过Tail-PCR鉴定转基因系中的基因组插入位点(图S6c,d)。
在5%次氯酸钠中灭菌并在水中浸泡12小时后胚根突破种皮,将WT和TaMPK3 -OE小麦植物的种子置于含有0、1、5或10μM ABA的湿滤纸上。所有种子在 ABA 处理的湿滤纸上在 12 小时内发芽,WT 和TaMPK3 -OE 小麦之间的种子发芽没有明显差异。然而,TaMPK3 -OE 小麦品系在不同浓度的 ABA 下表现出其生长表型的变异性(图S7a-c)。特别是,在 ABA 处理下, TaMPK3 -OE 幼苗的枝条长度显着长于 WT 的枝条长度(图S7d)。表型分析进一步表明,与 WT 相比,所有三个TaMPK3- OE 系在幼苗期都表现出降低的 ABA 敏感性(图 3)。更具体地说,在 ABA 处理 9 天后,TaMPK3 -OE 幼苗的枝条和根长显着长于 WT。此外,在对照条件下,在TaMPK3 -OE 和 WT 小麦之间未检测到幼苗茎长或根长的显着差异(图3a,b)。TaMPK3 -RNAi 系在幼苗期 ABA 处理的第 9天表现出不同的生长模式(图3c,d)。这些结果表明TaMPK3过表达减弱,TaMPK3-RNAi增强了ABA对小麦生长的抑制作用。

编辑切换为居中
添加图片注释,不超过 140 字(可选)
Fig. 3
3.4 ABA减少了TaMPK3和TaPYL4之间的相互作用,从而抑制了TaPYL4的降解
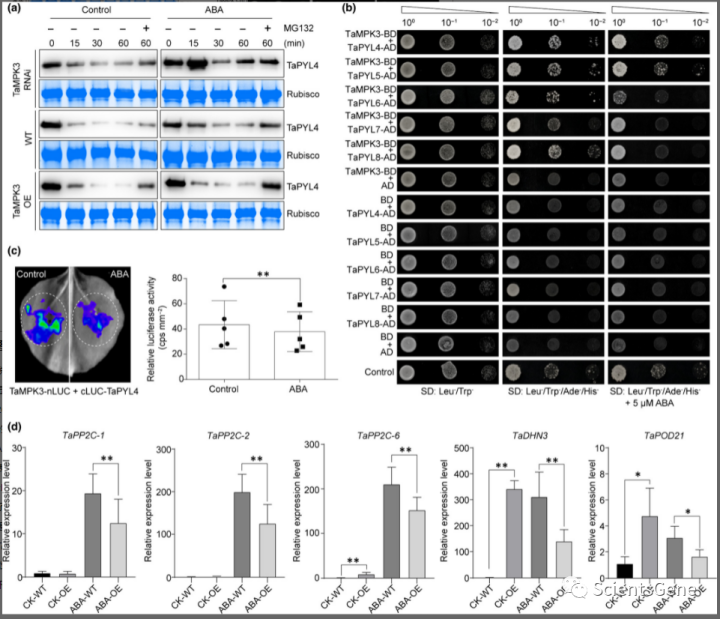
考虑到据报道TaPYL4过表达增加了小麦的 ABA 敏感性和耐旱性,而TaMPK3过表达导致对 ABA 的敏感性降低,因此我们假设 TaMPK3 可能会降低 TaPYL 的稳定性。无细胞裂解物的蛋白质印迹分析表明,TaPYL4 蛋白降解在TaMPK3 -RNAi 植物中受到显着抑制,但在TaMPK3 -OE 植物中与 WT 相比升高。此外,用 26S 蛋白酶体抑制剂 MG132 处理导致 TaPYL4 降解的部分抑制(图 4a),表明 TaMPK3 在调节小麦中 TaPYLs 的降解和稳定性中起主要作用。
由于 ABA 处理减少了TaMPK3 -OE、WT 和TaMPK3 -RNAi 小麦植物中 TaPYL4 的降解(图4a),因此我们试图确定 ABA 是否对 TaMPK3 和 TaPYL 之间的相互作用有任何影响。使用 Y2H 分析,我们证实暴露于 5 μM ABA 减少了酵母中 TaMPK3 和 TaPYLs 之间的相互作用,这表明与未处理细胞相比,与 TaMPK3-BD 和 TaPYLs-AD 共转化的酵母细胞生长较慢(图4b )。在烟草中,与对照叶片相比,我们还检测到在 10 μM ABA 处理下与 TaMPK3-nLUC 和 TaPYL4-cLUC 构建体共同浸润的叶片中的发光信号低于对照叶片(图4c)。YB2-nLUC 和 YA16-cLUC 用作阳性对照(图S8),其相互作用不受ABA处理的影响。与对 ABA 敏感性降低一致,随后基于 qPCR 的 ABA 信号通路基因表达(即TaPP2C-1、TaPP2C-2、TaPP2C-6、TaDHN3和TaPOD21)在 ABA 处理或 ABA 未处理的TaMPK3 -OE 和与 ABA 处理下的 WT 相比, WT 幼苗在TaMPK3 -OE 植物中的表达显着降低。此外,TaPP2C-6、TaDHN3和TaPOD21在未处理条件下,在TaMPK3 -OE 小麦中表现出显着更高的表达(图4d)。这些结果表明,TaMPK3 通过泛素化依赖性抑制 TaPYL4 有助于降低对 ABA 的敏感性。
编辑切换为居中
添加图片注释,不超过 140 字(可选)
Fig. 4
3.5 TaMPK3 可以以自磷酸化依赖性方式磷酸化 TaPYL4 并促进 TaPYL4在体外的降解
先前的研究表明,TaMPK3 可以使其 Tyr-196 残基自磷酸化,但不能使其 Ser 或 Thr 残基自磷酸化,并且它表现出自磷酸化依赖性蛋白激酶活性(Takezawa,1999)。为了评估其自磷酸化,我们首先生成了 TaMPK3 (TaMPK3 K65R ) 的变体,其中 65 位的赖氨酸 (K) 残基被转化为精氨酸 (R) 以消除其自磷酸化活性。然后我们培养纯化的 TaMPK3-GST、TaMPK3 K65R-GST 或 GST(空载体)蛋白与激酶反应缓冲液和 ATP,然后使用抗 GST 单克隆抗体通过 75 μM phos-tag™ 丙烯酰胺 AAL-107 测定法定量自磷酸化。结果显示WT激酶(TaMPK3-GST)经历了自磷酸化,由滞后迁移表明,而突变体TaMPK3 K65R -GST和单独的GST标签没有(图 5a)。

编辑切换为居中
添加图片注释,不超过 140 字(可选)
Fig. 5
3.6 外源 ABA 和干旱诱导的 TaMPK3 转录和蛋白质积累
据报道, TaMPK3的表达,也称为WCK-1,是由真菌诱导剂钙离子载体 A23187 以及干旱诱导的。在本研究中,我们进行了基于 qPCR 的 mRNA 定量和免疫印迹分析,以检查在 PEG6000 诱导的干旱或外源 ABA 处理下不同组织和幼苗期的TaMPK3表达模式。qPCR 测定的结果表明 TaMPK3 在营养组织中表达最高,尤其是在叶子中(图S11a),在 PEG 和 ABA 处理后,mRNA 水平增加,分别在叶子中的 0.5 和 1 小时达到峰值(图S11b)。与未处理的对照(图S11c)相比,外源 ABA(图S11d)和 PEG(图S11e)处理增加了 TaMPK3 蛋白的丰度,并且抗 Erk1/2 抗体用于蛋白质印迹(WB)测定(图S12 )。总之,这些结果表明 TaMPK3 表达在转录物和蛋白质水平上通过暴露于干旱条件或外源 ABA 上调。
3.7 小麦幼苗的耐旱性因TaMPK3过表达而降低,而因TaMPK3抑制而增加
根据我们对 TaMPK3 对 ABA 传感影响的发现,我们接下来研究了它的过表达或抑制如何影响小麦幼苗的耐旱性。为此,我们检查了TaMPK3 -OE、WT 和TaMPK3 -RNAi 幼苗的表型,这些幼苗在充分浇水的条件下发芽,但随后停止灌溉 18 天。值得注意的是,过表达系在发芽后第 12 天开始枯萎,到第 15 天变得松弛,而 WT 植物在第 13 天枯萎并在第 15 天变得松弛(图 6a)。发芽后第 12 天脯氨酸和第 15 天丙二醛 (MDA) 的测量表明, TaMPK3-中脯氨酸含量显着降低。OE植物比WT植物,而MDA含量显着更高(图6b,c)。在完全灌溉的 3 天恢复期后,我们发现TaMPK3 -OE 植物严重脱水并且基本上无法恢复(图6a),其存活率和枝条鲜重明显低于 WT(图6d,e) .

编辑切换为居中
添加图片注释,不超过 140 字(可选)
Fig. 6
相比之下,TaMPK3-RNAi植物在发芽后第 13 天开始枯萎,到第 16 天变得松弛(图6a)。然而,这些植物表现出比WT更耐旱的表型,具有更高的存活率、芽鲜重和脯氨酸含量,但与WT相比,MDA含量更低(图6b,d,e)。值得注意的是,WT 和TaMPK3 -RNAi 小麦幼苗在干旱处理后 3 天恢复后表现出相似的存活率 (> 80%),而TaMPK3 -OE 系的平均存活率为 3%。
3.8 TaMPK3负调控小麦营养生长期抗旱性
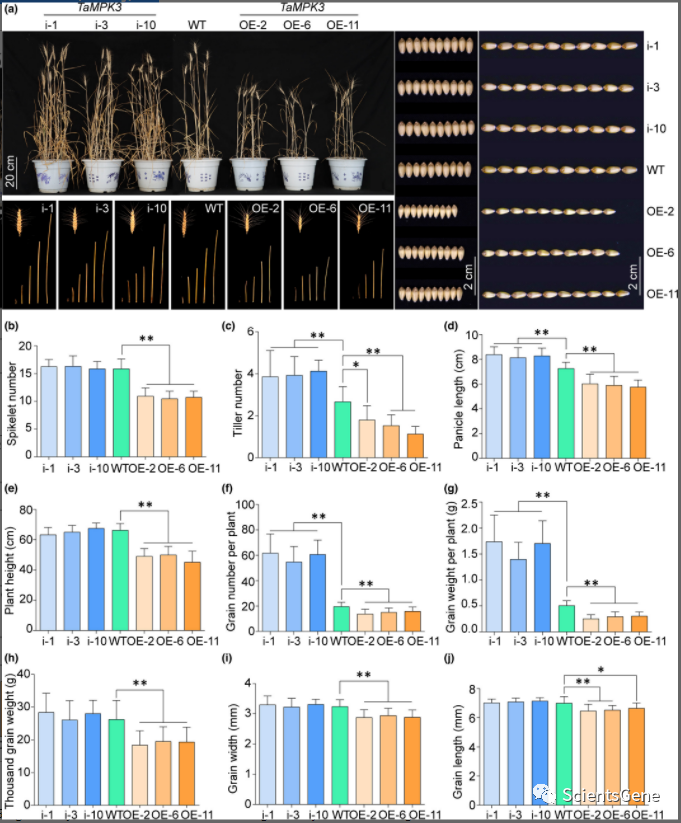
考虑到小麦幼苗中的这些影响,我们接下来检查了温室条件下成年(拔节)阶段TaMPK3 -OE、WT 和TaMPK3 -RNAi 植物的耐旱性和农艺性状表型。除了TaMPK3 -OE在水分充足的条件下比WT和TaMPK3 -RNAi小麦植株的株高短外,它们之间没有显着的表型差异(图S14)。在抽穗期停水 15 d,然后充分灌溉直至成熟后,我们观察到TaMPK3 -OE、WT 和TaMPK3 -RNAi 植物在干旱条件下的植物生长、穗和种子发育存在显着差异(图 7a),TaMPK3过表达对农艺性状产生明显的负面影响。例如,小穗数、分蘖数、穗长、株高、单株粒数、单株粒重、1000粒重、粒宽和粒长(图7b-j)在OE系中均显着降低。与干旱处理后的WT。相比之下,与WT相比,TaMPK3-RNAi植物表现出更高的分蘖数(图7c)、更长的穗长(图7d)、更高的每株粒数(图7f)和更高的每株粒重(图7g)。然而, TaMPK3之间没有发现显着差异-RNAi 和 WT 植物的小穗数(图7b)、株高(图7e)、1000 粒重、粒宽或粒长(图7h-j)。
编辑切换为居中
添加图片注释,不超过 140 字(可选)
Fig. 7
然后我们在田间条件下研究了TaMPK3- OE、WT和TaMPK3-RNAi小麦植株在开花期的耐旱性,发现结果与在幼苗(图6)和拔节期进行的耐旱性测定结果一致 (图7 ) 。在田间,TaMPK3 -OE、WT 和TaMPK3 -RNAi 小麦植株在水分充足的条件下没有显示出显着的表型差异,除了TaMPK3 -OE 植株的株高比 WT 和TaMPK3 -RNAi 株系短(图S15 )。然而,当开花后15 d停水时,TaMPK3- OE小麦植株的叶片变黄的时间比WT的要早,而WT的叶片变黄的时间又比TaMPK3-RNAi的稍早(图 8a)。值得注意的是,在田间干旱条件下, TaMPK3 - OE、WT和TaMPK3-之间的分蘖数、有效分蘖数、单株粒数、穗长、小穗数或粒长(图8) 与RNAi 小麦植物没有显着差异。随着干旱处理持续时间的延长,TaMPK3 -OE 品系最终表现出较低的千粒重(图8j)和粒宽(图 8jk ) 与 WT 和TaMPK3 -RNAi 植物相比。在田间干旱条件下, TaMPK3-RNAi小麦品系相对于WT的农艺性状没有显示出显着差异(图8)。这些结果表明,TaMPK3过表达对干旱耐受性和产量产生负面影响,而TaMPK3抑制导致积极或中性影响,具体取决于生长阶段和条件。

编辑切换为居中
添加图片注释,不超过 140 字(可选)
04
结论
随着周围环境恢复到非胁迫条件,植物应进行适当调整以发挥最佳功能。基于这一合理假设并结合我们的实验结果,我们提出 TaMPK3 抑制 TaPYL 活性以削弱 ABA 信号传导是抑制干旱后恢复过程中的应激反应和促进正常生长途径激活的关键步骤。
考虑到TaMPK3-RNAi可以显着提高小麦的耐旱性,而不会明显影响其他农艺性状,我们提出调节TaMPK3的表达可以通过减弱其对ABA信号的抑制来提高植物的耐旱性。这种潜在的应用对于专注于作物胁迫耐受性的育种计划很有前景,并为多种生长条件下的 TaMPK3 表达提供有价值的参考数据。
然而,目前尚不清楚是什么触发了干旱后恢复下 TaMPK3 的高表达。总的来说,我们的研究揭示了正常、干旱和干旱后恢复期的动态分子调节机制,通过 TaMPK3 介导的通过 TaPYL 的相互作用和靶向降解抑制 ABA 信号传导(图 9)。

编辑切换为居中
添加图片注释,不超过 140 字(可选)
05
获取原文
原文链接:
https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.18326
赛思基因www.scientsgene.com有限公司致力于开发转基因和基因编辑中的“硬核科技”,突破遗传转化中的基因型限制,辅助基因编辑育种。目前公司拥有国内数量最多的稳定遗传转化体系,包括但不限于油茶、花生、藜麦、大白菜、小白菜、辣椒、菜心、油菜、紫菜薹、樱桃萝卜、胡萝卜、甘蓝、抱子甘蓝、宝塔菜、紫/黄/绿花椰菜、青花菜、西瓜、南瓜、甜瓜、茄子、梨树、枣树、大豆、棉花等遗传转化体系。
赛思基因可为广大科研和育种工作者提供稳定的,非嵌合的,多品种的过表达、基因沉默株系和基因编辑突变体。期待与您的深入合作,共同推动中国种业的进步。