–https://doi.org/10.1038/s41590-024-01901-1
BACH2 regulates diversification of regulatory and proinflammatory chromatin states in TH17 cells
留意更多内容,欢迎关注微信公众号:组学之心
研究团队和研究单位
Aviv Regev–Genentech

Vijay K. Kuchroo–Broad Institute of MIT and Harvard–Klarman Cell Observatory

研究简介
研究背景
-
平衡促炎和调节性 CD4+ 辅助 T 细胞亚群的重要性:既可以抵抗病原体和肿瘤,还能避免不受控制的炎症和自身免疫。
-
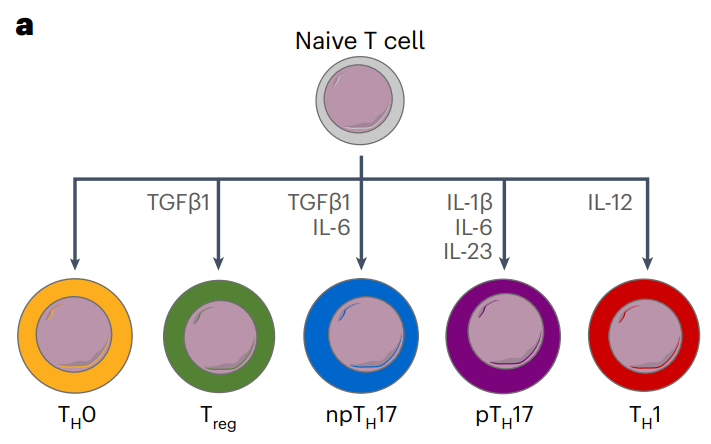
TH17 细胞的双重角色:TH17 细胞通过产生 IL-17 在宿主防御细胞外病原体和维持粘膜屏障稳态中起关键作用,但它们也是自身免疫性疾病的重要致病因素。非致病性 TH17 (npTH17) 细胞在体外由 IL-6 和 TGFβ 分化,几乎不引发组织炎症。致病性 TH17 (pTH17) 细胞由 IL-6、IL-1β 和 IL-23 共同作用产生。
-
TH17 细胞的功能和可塑性:TH17 细胞表现出额外的可塑性,能够获得与其他 CD4+ 辅助 T 细胞亚群相关的功能或致病促炎状态。例如,在自身免疫模型中,TH17 细胞可以转变为 TH1 样细胞,从而在特定疾病模型中引发病理变化。
研究内容
-
调控途径:研究发现诱导 TH17 异质性的调节途径,并发现 npTH17 和 pTH17 细胞在染色质景观上存在显著差异。npTH17 细胞与调节性 T 细胞共享染色质配置,而 pTH17 细胞则表现出 npTH17 和 TH1 细胞的特征。
-
单细胞技术的应用:结合单细胞转座酶可及染色质测序 (scATAC-seq) 和单细胞 RNA 测序 (scRNA-seq),研究推断出控制不同细胞状态的调控网络。此外,将 TH17 致病性的染色质特征映射到效应基因,并预测 pTH17 细胞程序的多个转录因子调控因子。
-
BACH2 的作用:研究证实 BACH2 在体外和体内促进免疫调节 npTH17 程序并抑制 TH17 细胞中的促炎性 TH1 样程序。
总的来说,研究确定了 TH17 异质性的调控因子,这些因子可能成为减轻自身免疫的潜在靶点,有助于设计专门针对致病 TH17 群体的治疗策略。
研究结果
1.npTH17 和 pTH17 细胞具有不同的染色质图谱
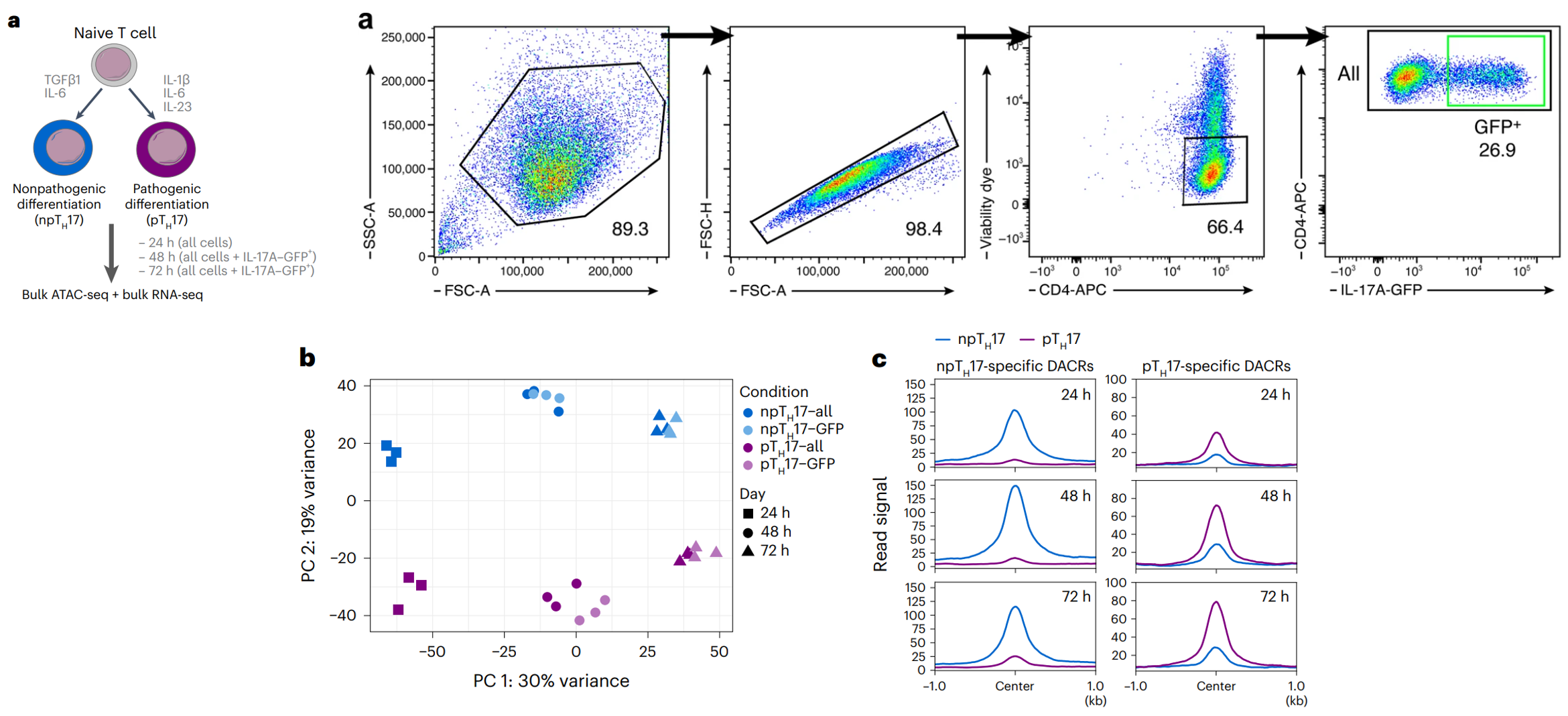
(a左上)研究诱导刺激幼稚辅助性 T 细胞 24、48 和 72 小时后,对体外分化的 npTH17 和 pTH17 细胞进行了bulk ATAC-seq和bulk RNA-sq,获得这两种细胞的染色质可及性大体的情况。
(另一个a)为了评估 IL-17A 表达与染色质之间的关系,研究还从 IL-17A GFP 小鼠挑出 TH17 细胞后也做了测序,并与所有 CD4+ 细胞进行了比较。
(b)主成分分析表明 npTH17 和 pTH17 细胞之间染色质景观的显著差异,两种类型的 TH17 细胞至少在用细胞因子处理后 24 小时内就出现了分化,这证明它们是具有不同基因调控机制的不同细胞命运。
(c)展示了两个细胞亚群的差异染色质可及性区域(DACRs),48h的 pTH17 和 npTH17 细胞之间差异可及峰数量最多(7,039 个)。

(d)给DACRs注释后发现绝大多数位于非编码区(内含子Intron和基因间Intergenic),距离最近的转录起始位点(TSS)较远(>25,000 个碱基对 (bp)),突出表明远端调控元件是致病和非致病细胞状态的关键区别特征和潜在调控中心。
(e)在 48 小时检测到的DACR 中,有4,645 个在 npTH17 细胞中更易接近,2,394 个在 pTH17 细胞中更易接近。这些包括位于 TH17 效应基因近端位点的 DACR,例如 Il17a 和 Il17f(在 npTH17 细胞中较高)、与调节性 TH17 程序相关的基因(Il10在 npTH17 细胞中较高)和与促炎性 TH17 基因程序相关的基因(例如,Ifng在 pTH17 细胞中较高)。

(c-d)为了测试这些染色质可及性差异是否也反映在相应的人类细胞中,研究对体外培养的人类 IL-10− IFNγ+ 或 IL-10+ IFNγ– TH17 细胞进行 ATAC-seq 分析,这些细胞要么未受刺激(0 小时),要么在 6 或 72 小时时重新受刺激。PCA分析可见人类 IL-10− IFNγ+ 和 IL-10+ IFNγ– TH17 细胞之间染色质景观的显著差异,和小鼠细胞中观察到的染色质情况相对应。具体而言,PC1 反映了 IL-10− IFNγ+ 和 IL-10+ IFNγ– TH17 细胞随时间的变化相似,而 PC3 在每个时间点将 IL-10– IFNγ+ 与 IL-10+ IFNγ– TH17 细胞分开。
(e)IL-10– IFNγ+ 细胞的峰与促炎基因(BHLHE40、CD69、IL6 和 LGALS3)相关,而 IL-10+ IFNγ– 细胞中的开放峰与调节基因以及与小鼠 npTH17 细胞相关的基因(AHR、BATF、CCR6、IL17A、IL17F、MAF 和 NRP1)相关。
表明具有不同功能的人类 TH17 细胞也代表具有不同基因调控机制的不同细胞命运。
2.染色质可及性变化与 TH17 细胞中的基因表达差异相对应
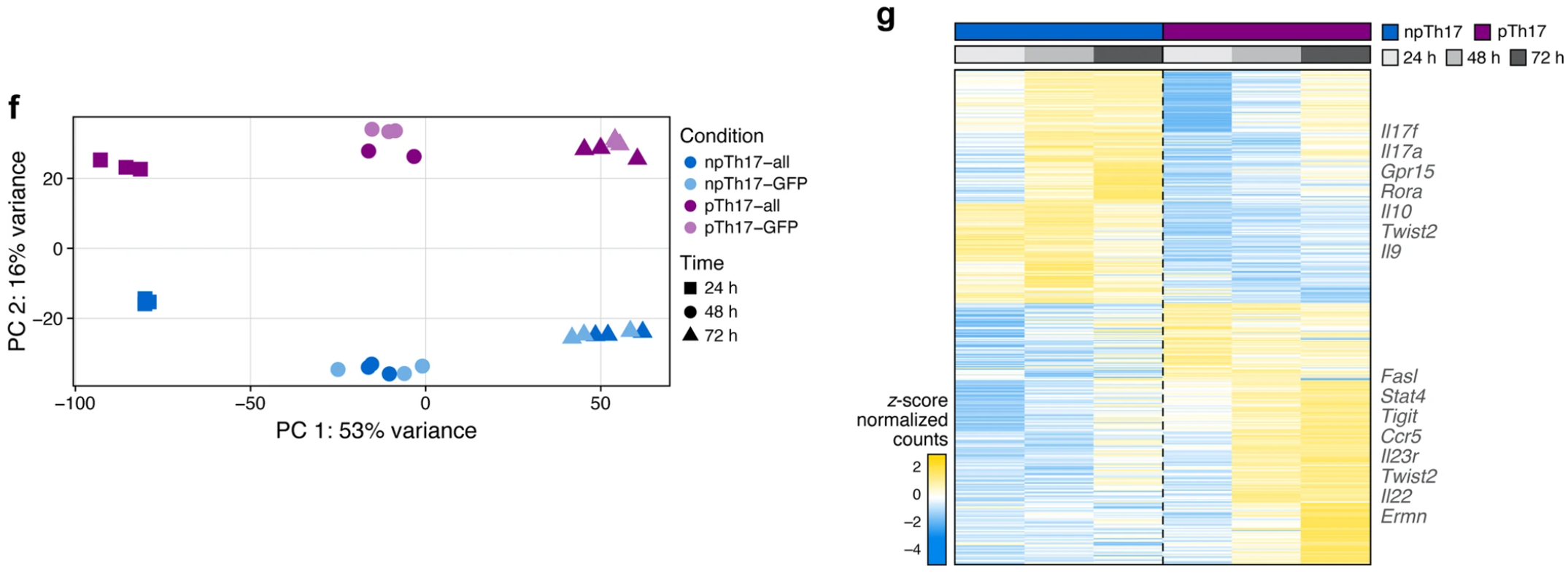
体外极化的 TH17 细胞群的 RNA-seq 数据大体如图所示,PCA分析可以看出两种细胞的转录水平差异很大,而且按照时序排布,表明至少在24小时后功能上已经有较显著的差别。

npTH17 和 pTH17 细胞在染色质景观中的差异、与基因表达程序的差异之间存在很强的正相关性。在 416 个差异表达基因中(取每个时间点按Fold change 排名的前 200 个,FDR < 0.05),240 个具有附近的 DACR,且具有显着的正相关性(平均 Pearson r = 0.70,FDR < 0.05)。
与染色质变化相关的 npTH17 特异性基因包括免疫调节基因 (Il9、Maf、Ahr、Cd5l 和 Il10)以及尚未与 npTH17 细胞相关但在其他免疫细胞类型中具有先前定义的调节功能的基因。pTH17 特异性基因包括已确定的促炎基因 (Il23r、Stat4、Batf3、Gzmb 和 Nkg7) 和先前未涉及 pTH17 表型或功能的基因 (Il33 和 Ermn)。此外,几个染色质可及性发生变化的关键基因座 (例如,Ifng 和 Csf2) 没有相应的基因表达差异,可能是这些基因座中的基因在允许的染色质环境下做好转录的准备。
总体而言,这些结果表明 pTH17 和 npTH17 细胞具有与促炎和调节基因程序相关的不同表观遗传调控机制。
3.在实验性自身免疫性脑脊髓炎(EAE)期间,引流淋巴结 (dLN) 和中枢神经系统 (CNS) 的 TH17 细胞染色质景观

为了探究 npTH17 和 pTH17 细胞的染色质景观与疾病的关系,研究收集了 EAE 小鼠(多发性硬化症 (MS) 的动物模型)的 TH17 细胞的 ATAC-seq。已有 scRNA-seq 研究证明 EAE 小鼠中体外分化的 npTH17 细胞与 pTH17 细胞的细胞状态,以及 dLN 衍生的 TH17 细胞与 CNS 衍生的 TH17 细胞的细胞状态之间存在相似性。因此,本研究进一步探索在 EAE 期间, dLN 和 CNS 中 TH17 细胞的染色质可及性与疾病状态的联系,在疾病危险期从 dLN 和 CNS 中分选出存活的 IL-17A–GFP+ CD4+ T 细胞,并进行bulk ATAC-seq 和 RNA-seq。
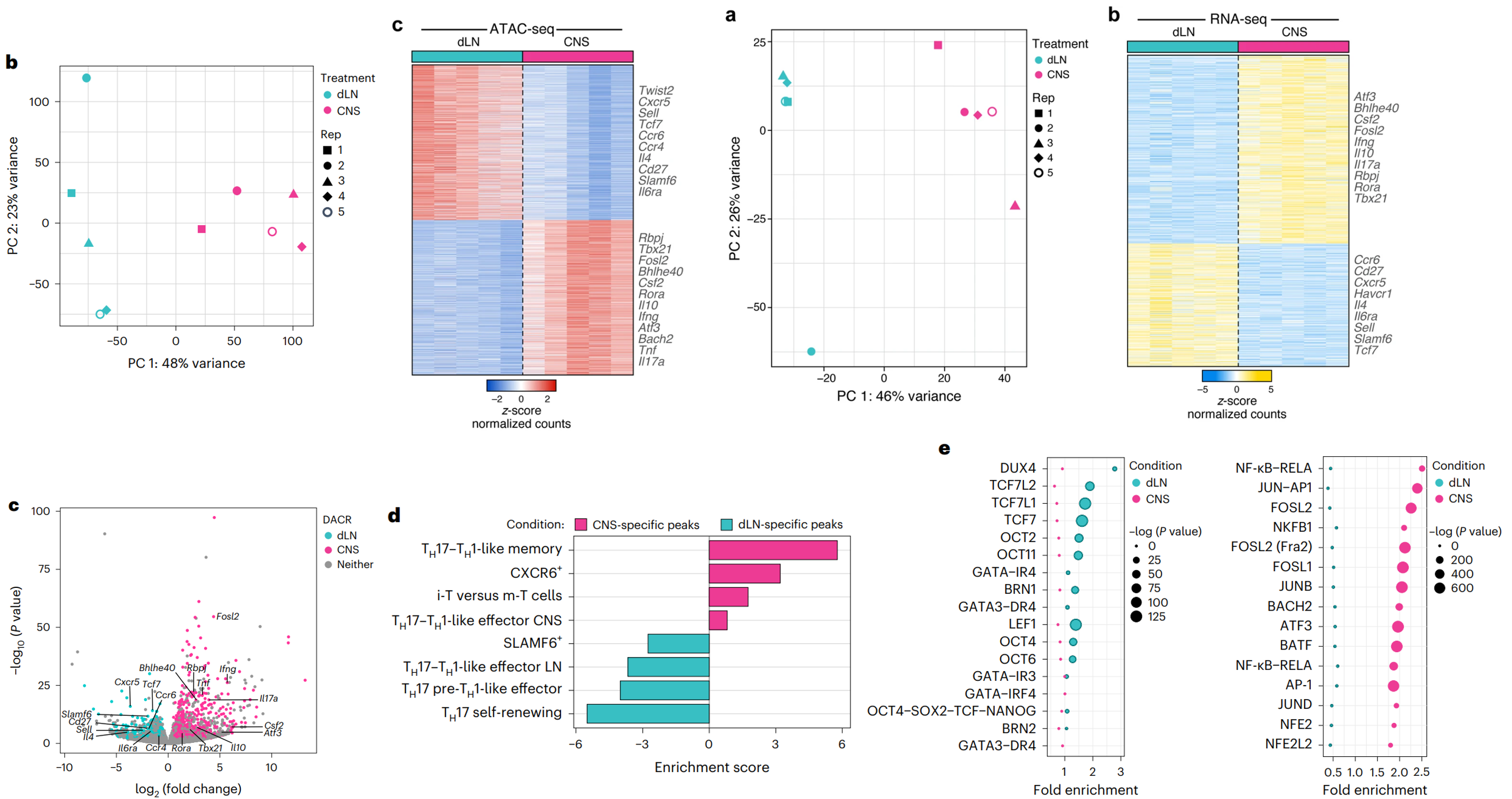
(上半部分+下半部分的c)dLN 和 CNS 中 TH17 细胞的染色质可及性和基因表达水平存在显着差异。dLN 特异性峰接近与 npTH17 表型相关的基因 (Cxcr5、Sell、Tcf7、Il6ra 和 Maf)。此外,Il4 基因座在在 dLN TH17 细胞中更容易接近,并且该基因在该细胞中的表达更高。表明dLN TH17 细胞倾向于表达 IL-4 并可能转换为 2 型辅助 T (TH2) 细胞表型(IL-4是TH2的biomarker)。CNS 特异性峰和 TH17 致病性的基因位点 (Rbpj、Tbx21、Bhlhe40、Csf2、Ifng 和 Il17a) 有联系,以及先前未定义的基因位点,如 Bach2。
(d)CNS TH17 细胞具有与 TH17-TH1 细胞相似的特征,而 dLN TH17 细胞具有自我更新的干细胞样 TH17 细胞特征。
(e)dLN 和 CNS 的 TH17 细胞的 DACR 富集了 npTH17 和 pTH17 细胞的调节因子的motifs。dLN 特异性峰富集了与类干细胞和类自我更新 TH17 程序相关的 TF 位点,如 TCF1、LEF1 和 OCT4;而 CNS 特异性峰富集了已知在 TH17 分化和致病性中发挥作用的 TF 位点,如 FOSL2、BATF 和 JUNB。
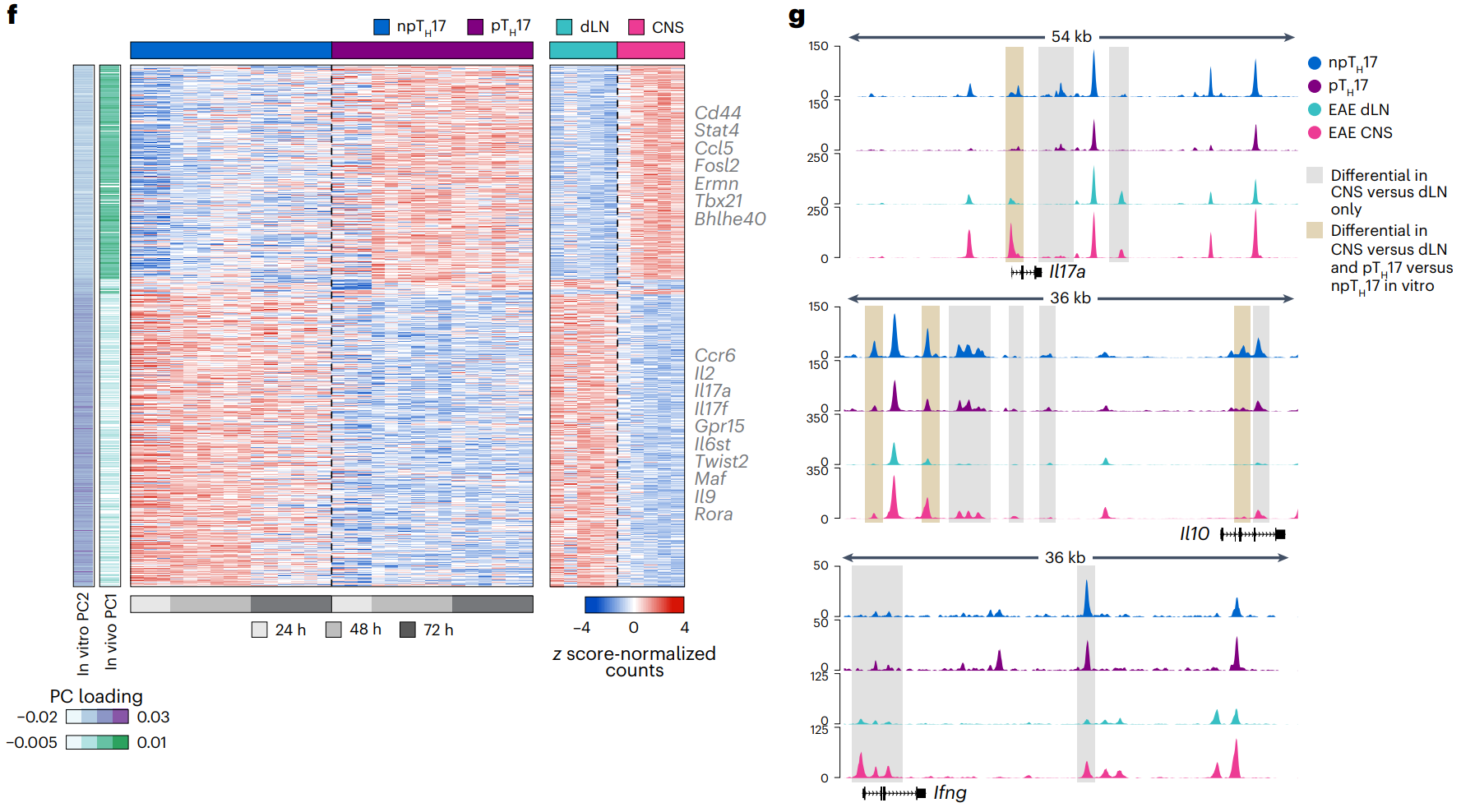
将体外极化细胞和体内衍生细胞的染色质特征进行比较后,确定了 491 个峰在 npTH17 和 dLN 细胞之间具有共同的差异可及性区域,678 个峰在 pTH17 和 CNS 细胞之间具有共同的差异可及性区域。
CNS TH17 细胞和 pTH17 细胞中具有共同差异可及性的峰,有参与 TH17 激活和致病性的位点(Cd44、Stat4、Fosl2、Tbx21 和 Bhlhe40)。 dLN 衍生细胞和 npTH17 细胞中差异可及峰有 npTH17 细胞的特征位点(Ccr6、Il6st、Maf 和 Il9)以及先前未定义的候选位点(Gpr15 和 Twist2;FDR < 0.05)。这些峰与体外和体内TAC-seq 谱的相应 PC 分数一致,高 PC2 主要出现在 npTH17 细胞和 dLN TH17 细胞中,高 PC1 主要出现在 pTH17 细胞和 CNS TH17 细胞中。
表明,pTH17 细胞和 CNS TH17 细胞与 TH17 细胞的致病性功能相关,而 npTH17 细胞和 dLN TH17 细胞则与非致病性或调节性功能相关。
4.TH17 细胞与其他 CD4+ T 细胞的染色质特征
TH17 细胞与其他 T 细胞亚群之间的关系尚未得到充分的探索。为了更全面地了解 TH17 细胞的可塑性,研究比较了体外衍生的 npTH17、pTH17、TH1、Treg 细胞和无细胞因子分化的活化 T 细胞 (TH0) 之间的 ATAC-seq 和 RNA-seq。

CD4+ T 细胞亚群之间的染色质可及性存在很大差异,每个亚群根据前两个 PC 分别分组,形成两个主要“分支”,一个从 TH0 到 TH1、pTH17 和 npTH17 细胞,另一个到 Treg 细胞。根据 PCA 和成对相关性确定,Treg 细胞最为独特。尽管在 PC1 上Treg 细胞和 npTH17 细胞距离最远,但它们在 PC2 上距离最近,表明存在一组潜在的共同特征。这也反映在它们的 RNA-seq 图谱中(PC2),可能因为它们分化都需要 TGFβ。pTH17 细胞定位在 npTH17 和 TH1 细胞之间,并且在染色质谱中与 TH1 细胞最相关。
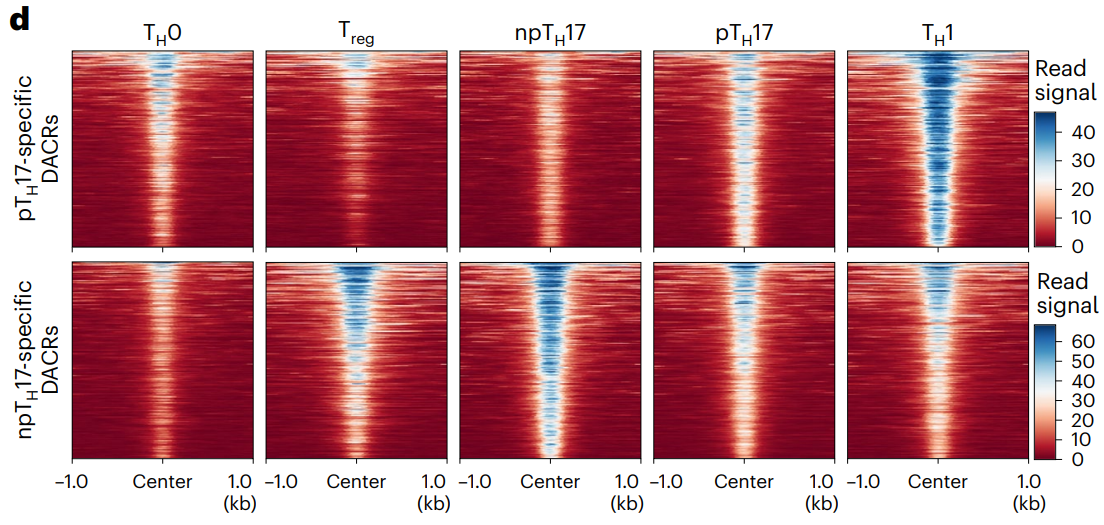
pTH17 细胞中比 npTH17 细胞中更开放的 DACR 在 TH1 细胞中显示最高信号。在 Treg 细胞中检测到 npTH17 特异性 DACR,支持促炎和稳态 TH17 细胞的双重功能细胞状态反映了 TH17 细胞激活其他 CD4+ T 细胞类型特征程序的能力。

(e)对从 npTH17、pTH17、TH1 和 Treg 细胞的成对比较中确定的 DACR 进行了聚类(k 均值聚类,k = 10)。簇 1 由所有效应 CD4+ 辅助 T 细胞共享。簇 2 和簇 3 主要由 Treg 细胞特异性峰组成,其中一些峰接近对 Treg 细胞功能很重要的基因,例如 Nrp1、Il10rb1 和 Irf1。簇 4 具有 npTH17 和 Treg 细胞之间共享的 DACR。这些 DACR 包括那些靠近先前与 TH17 和 Treg 细胞中的功能相关的基因(Il9、Gpr15 和 Il2ra)。簇 5 和簇 6 包含 npTH17 细胞特有的峰。簇 7 包括 npTH17 和 pTH17 细胞之间共享的峰,并且特定于一般的 TH17 细胞表型。簇 8 突出显示了区分 pTH17 和 npTH17 细胞的峰,这些峰在 TH1 细胞中也存在。簇 10 特定于 TH1 细胞,包括接近 TH1 细胞标记基因的峰,例如 Stat1、Il12rb2、Nkg7、Ifng 和 Plac8。
(f,a,k)根据染色质可及性预测与每个 DACR 相关的基因,并检查选定基因的表达,表征不同T细胞类型的转录水平和染色质可及性。

为了测试体外分化的 npTH17 细胞和 pTH17 细胞是否可以转分化为 TH1 细胞和 Treg 细胞,研究进行了体外转分化实验。尽管 npTH17 细胞中的 IFNγ 表达保持不变,但 pTH17 细胞在 IL-12 处理后显著上调了 IFNγ 的表达,这表明体外分化的 pTH17 细胞比 npTH17 细胞更容易呈现 TH1 细胞的特征。添加 TGFβ 会下调 pTH17 细胞中 FoxP3 或 IL-10 的表达,而不会改变 npTH17 条件下 FoxP3 或 IL-10 的表达,可能是因为 TGFβ 信号在 npTH17 细胞极化中已经活跃。
5.pTH17 细胞维持在 npTH17 和 TH1 细胞之间的中间状态

为了检查 48 小时时染色质和表达如何随时间变化,研究在 TH1、pTH17 和 npTH17 细胞体外极化 0、1、6、12、20 和 48 小时期间进行了bulk ATAC-seq 和 RNA-seq。
PCA 确定了三个主要变异轴:“时间”(PC1),其中三种类型的样本进展相似,“激活”(PC2),其中所有类型从 1 小时增加到 6 小时,以及“极化条件”(PC3),其中来自不同条件的样本随着时间的推移变得越来越不同。具体而言,pTH17 和 npTH17 细胞的样本首先与 TH1 细胞变得不同(6 小时后),然后 npTH17 样本与 pTH17 细胞的区别越来越明显,以至于到 48 小时后,pTH17 细胞就成为 TH1 和 npTH17 细胞之间的中间体。在 ATAC-seq 和 RNA-seq 数据中观察到的相似模式,表明染色质和基因表达变化在 T 细胞极化过程中是协调一致的。

(左c)染色质区域在不同条件下随时间变化的可及性存在差异,可分为 10 个簇(k 均值聚类,n = 10,5,202 个峰),包括细胞类型特异性簇(簇 3、4、5 和 9)以及 pTH17 和 npTH17 细胞之间共同的簇(簇 2、7 和 8)、pTH17 和 TH1 细胞之间共同的簇(簇 6)或所有条件下共同的簇(簇 1)。在 5,202 个top DACR 中,52%的也通过从 PC1、PC2 或 PC3 加载进入了前 20% 的峰值。大多数(2,091 个峰,77%) 与 PC3 共享,特别是在簇 4、5、7、8、9 和 10 中,这与 PC3 对极化条件进行分层一致。
(d-右c)在 48 小时内在极化过程打开(d)或关闭(c)的 58,356 个和 60,452 个细胞类型特异性染色质可及性区域中,34% (19,739) 和 62% (37,204) 在不同条件之间共享。
6.单个 pTH17 细胞的分子谱介于 TH1 和 npTH17 细胞之间
为了区分 pTH17 细胞与 TH1 或 npTH17 细胞之间的共享和发散模式是否能够代表细胞内在的中间状态或细胞间异质性,研究接下来进行了基于scATAC-seq。
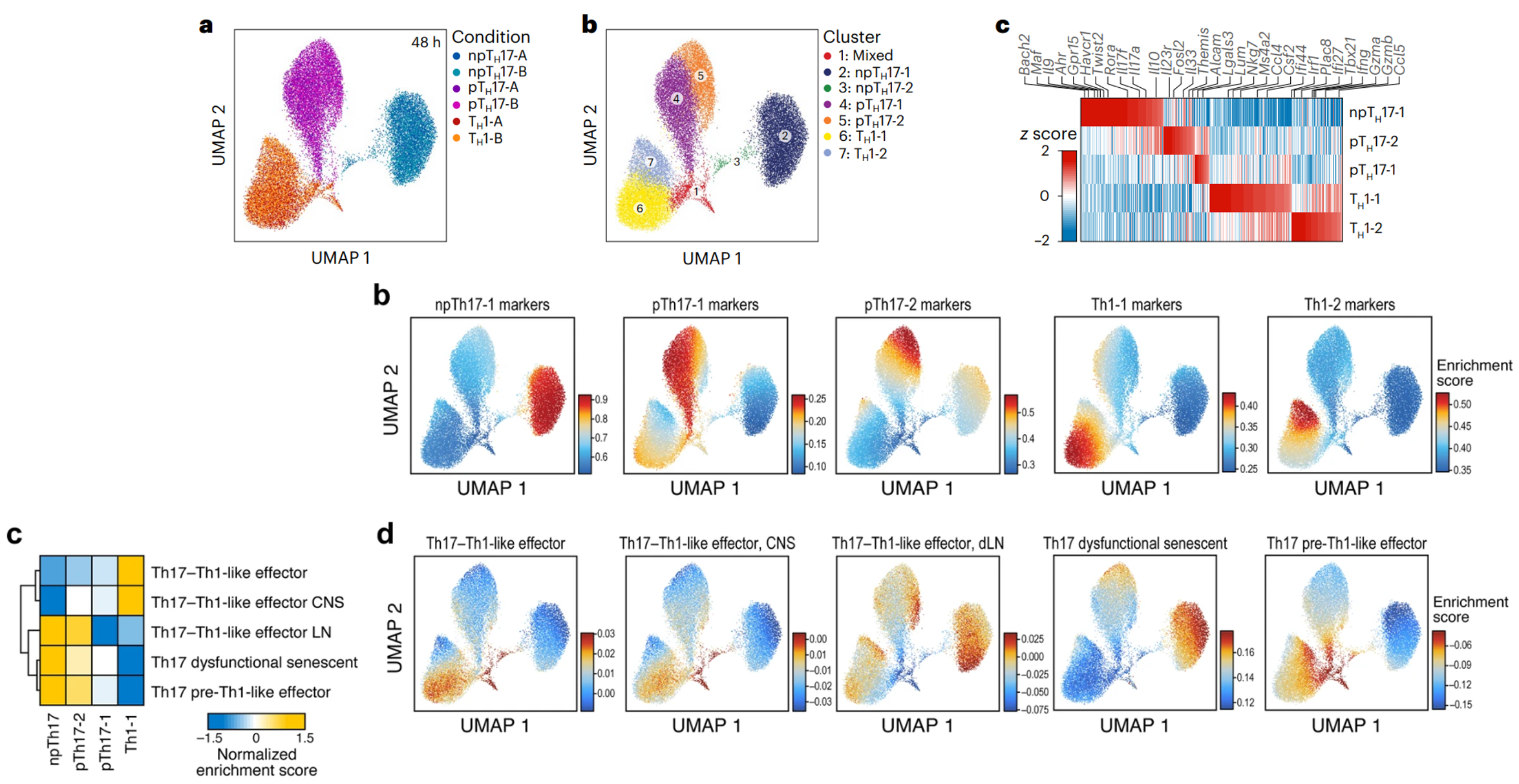
(上a-c)按细胞类型分组的 scATAC-seq 谱在很大程度上反映了在群体水平上观察到的 TH1 细胞到 pTH17 细胞到 npTH17 细胞的顺序,表明存在连续的细胞内在表型。划分为五个簇:一个 npTH17 细胞簇、两个 pTH17 细胞簇和两个 TH1 细胞簇。每个簇与不同的标记基因相关,与文章开头bulk ATAC-seq 为 npTH17(Il17a、Il17f、Twist2、Il10 和 Havcr1)、pTH17(Themis、Il33、Fosl2 和 Il23r)和 TH1(Csf2、Ccl5 和 Tbx21)细胞簇鉴定的基因一致。
(上b、c;下b)pTH17 细胞分布在 npTH17 和 TH1 细胞之前,且 pTH17 染色质可及性biomarker排布也位于它们之间。
(下c-d)pTH17 细胞的体内特征在 pTH17-1 和 TH1-1 簇中富集,而 npTH17 细胞的体内特征在 npTH17 和 pTH17-2 簇中富集。
可见pTH17 细胞的分子谱介于 TH1 和 npTH17 细胞之间。
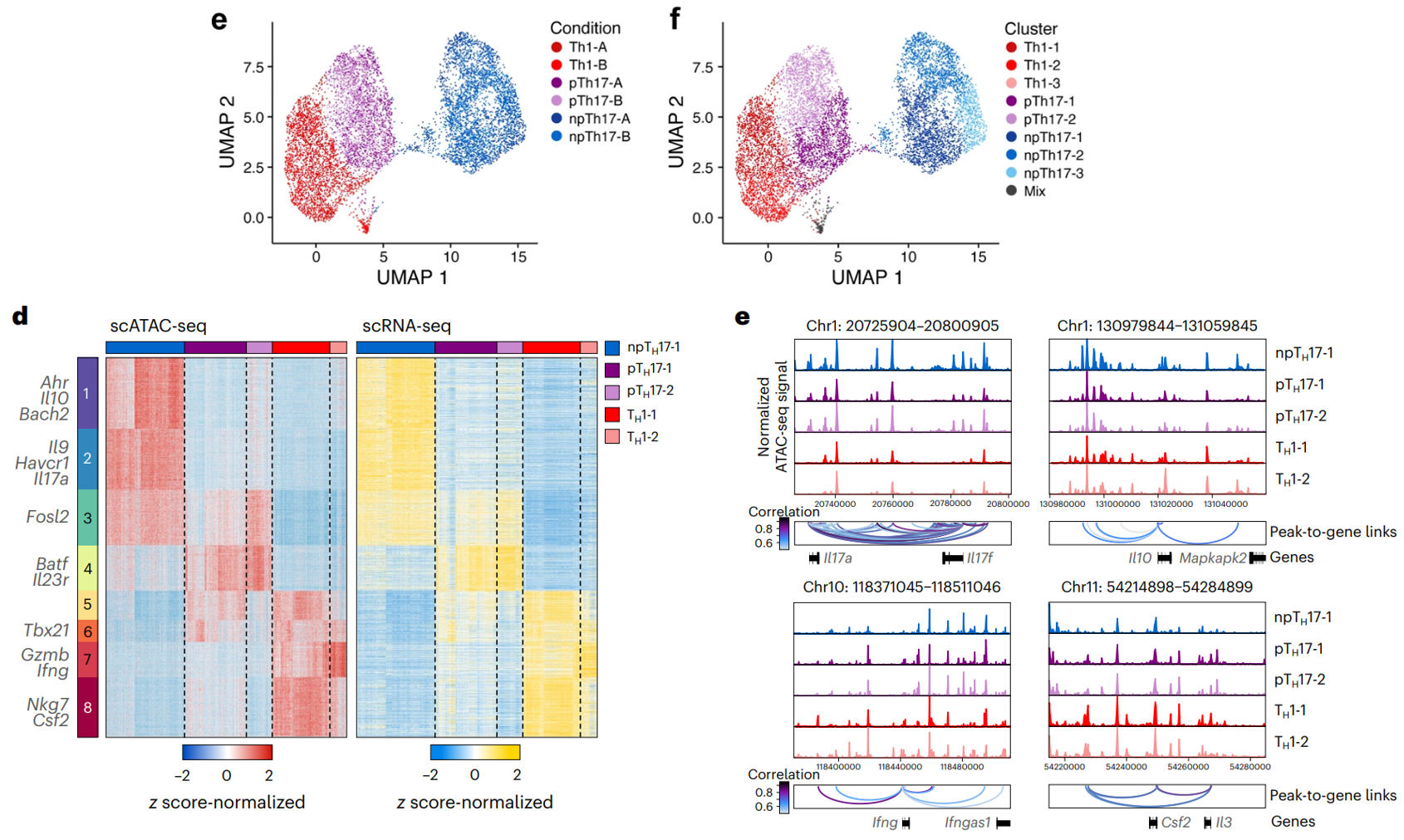
(e-f)将 scATAC-seq 数据与匹配的 scRNA-seq 谱整合在一起,利用它们将染色质峰可接近性与基因表达关联起来。
(d)23,608 个已识别的peak to gene(相关性 >0.45,FDR < 0.05)分为八个簇(k 均值聚类)。这些包括 npTH17 细胞的细胞类型特异性峰簇(簇 1 和 2;8,260 个 peak to gene,例如 Ahr、Il10、Bach2、Il9、Havcr1 和 Il17a)、pTH17 细胞(簇 4;2,827 个,例如 Batf 和 Il23r)和 TH1 细胞(簇 7 和 8;5,888 个,例如 Gzmb、Ifng、Nkg7 和 Csf2)以及 npTH17 和 pTH17 细胞共同的簇(簇 3;3,442 个,例如 Fosl2)和 pTH17 和 TH1 细胞共同的簇(簇 5、6 和 3;191 个,例如 Tbx21)。
此外,pTH17-2 细胞在与 npTH17 细胞共同峰的位点有较高的基因表达,而 pTH17-1 细胞和 TH1 细胞特异性基因的共同簇 5 和 6 中具有高基因表达和容易接近的染色质区域。
(e)peak to gene 包括关键的 TH17 细胞效应基因位点,例如 Il17a、Il17f、Il10、Csf2 和 Ifng,其峰与基因表达高度相关,可能作为潜在的调控元件。

为了进一步探索共享和细胞类型特异性的调控程序,研究对 scATAC-seq 数据的进行topic建模。细胞类型特异性topic以定义 npTH17 细胞的可接近染色质区域为特色,topic 11 区分了具有靠近致病性相关基因的可接近染色质区域的 pTH17 细胞,topic 13 代表具有与促炎基因程序相关的可接近染色质区域的 TH1 细胞程序。在捕获多种细胞类型内变异的topic中观察到可塑性,例如 pTH17-1 和 TH1-1 簇中的共享 npTH17 和 pTH17 细胞程序(topic 3)和共享 pTH17-1 和 TH1-1 程序(topic 9)。
总体而言,pTH17 细胞的范围很广,从 npTH17 细胞样特征更突出的细胞到具有更强的促炎 TH1 细胞样染色质状态的细胞。
7.TH17 和 TH1 细胞之间的调控网络,该网络可以预测 TH17 细胞致病性的调控因子
接下来,研究使用整合的 scATAC-seq 和 scRNA-seq 谱来预测结合 T 细胞类型特异性可及染色质位点的 TF 调控因子。
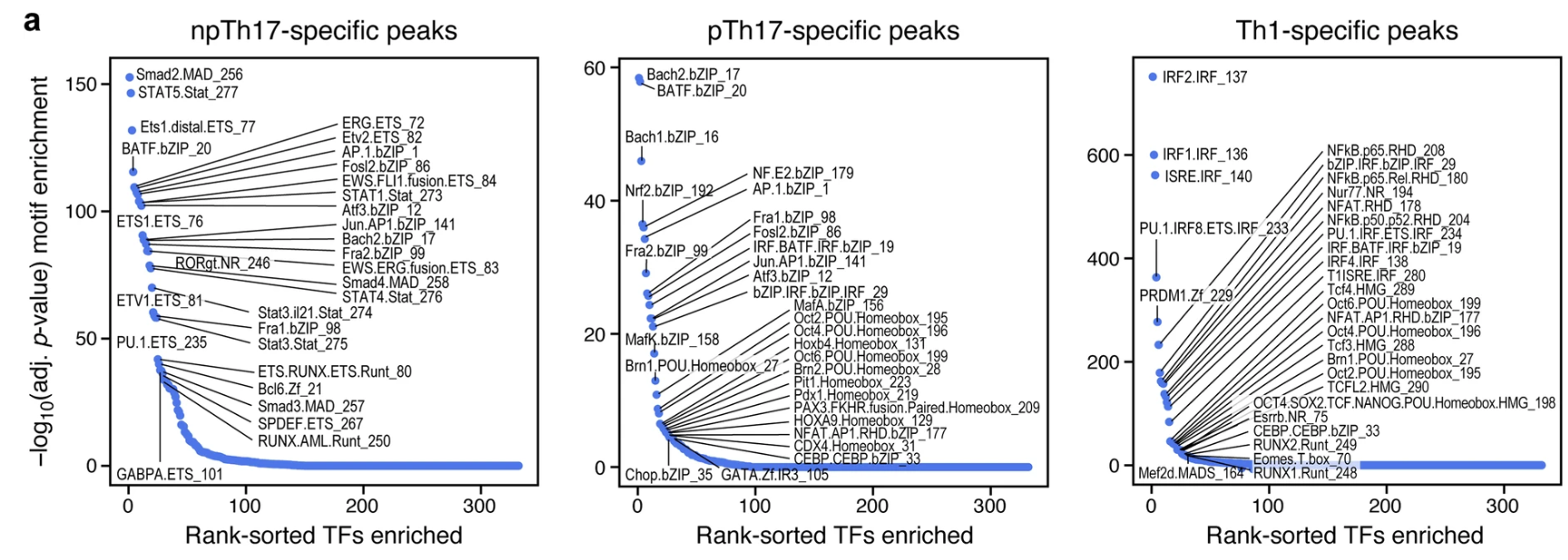


(a,g)a图展示了在 npTH17、pTH17 和 TH1 细胞特异性峰中富集的转录因子 (TF) motif。然后确定了 RNA 表达与基因座中含有同源motif的可及性变化区域呈正相关的 TF。该分析确认了编码 TH17 细胞调控因子的基因,如 Ets1、Junb、Fosl2 以及 Bach2。
(b)研究进一步构建 TF 靶标调控网络,通过从 scRNA-seq 数据中识别差异表达基因(在细胞亚群之间成对比较)并通过peak to gene连锁分析在与这些基因相关的峰中寻找 TF motif。如果在与基因相关的峰中发现 TF motif,并且 TF 和靶基因表达在 scRNA-seq 数据中的所有细胞中都相关,则得到的网络将 TF 与靶基因相关联。高度交互的 TF 调控因子(至少有十个靶标)和靶基因(至少有五个 TF)确定后,总计 78 个 TF、223 个靶基因和 3,749 条边。
(h,i)研究确定了一个针对代表性基因的网络,涵盖 24 个 TF 和 33 个目标基因,其中有 201 个激活边和 114 个抑制边。这是一个很大程度上自我强化、相互对抗的组织,因此,一种状态的基因程序的“激活因子”充当另一种状态的抑制因子。
Rorc、Ets1、Fosl2、Rbpj 和 Bach2 是调节 npTH17–pTH17 靶模块 1 和 2 的 TF 基因。Bach2 基因与 TH17 细胞基因调控无关,但研究预计是该模型中 TH17 细胞基因程序的主要驱动因素。
pTH17 细胞 TF 和基因主要位于与 npTH17 或 TH1 细胞共享的模块中。例如,pTH17–TH1 目标模块 3 和 5受 Tbx21、Eomes、Nr4a1 和 Atf3 正向调控。ATF3 是一种抗炎调节剂,可抑制巨噬细胞中的 Il6 和 Il12b 转录,并正向调节 CD4+ T 细胞中的 Il10 表达。然而,ATF3 也正向调节 TH1 细胞中的 Ifng 表达。

为了阐明 ATF3 在 T 细胞中的作用机制,研究进行了以 ATF3 为中心的子网络分析,预测 ATF3 激活致病的程序和抑制调节的程序。

结合第五部分的内容,研究发现48小时时候的网络分析鉴定出的许多调控因子,也是通过在沿极化时间进程变化的每个染色质区域簇的峰上富集TF motif 来鉴定的。例如,第 7 簇包括 TH17 细胞中常见的染色质区域,这些区域早期(6 小时)开放,有研究表明可能受已知的 TH17 细胞调控因子来调节,包括 BATF、JUNB 和 FOSL2,以及新型调节因子 ATF3 和 BACH2。
8.BACH2 作为 TH17 细胞致病性的负调控因子发挥作用
接下来就要对 BACH2 进行功能验证了,根据文章在此之前的测序数据分析结果,BACH2 很可能是 TH17 细胞染色质景观的一种新型调控因子。多种证据支持 BACH2 是 TH17 细胞状态的驱动因素:
- (1) 在 EAE 细胞的 TF motif 分析中发现了 BACH2;
- (2) npTH17 特异性峰与 Bach2 基因座接近;
- (3) Bach2 在 npTH17 细胞中表达水平较高;
- (4) BACH2 是 npTH17 细胞靶标的正调控因子;
- (5) 预测 BACH2 会随着时间的推移调节 TH17 细胞特异性染色质区域。

(左a)研究使用 CRISPR-Cas9 系统生成了 Bach2 敲除 (Bach2 KO) TH17 细胞,在 TH17 细胞中的 Bach2 基因座处形成了高效的插入/缺失 (indel),抑制BACH2基因的表达。
(右a)TH17 细胞分化过程中的 Bach2 KO 导致 npTH17 细胞中促炎细胞因子 IL-17A、IFNγ 和 GM-CSF 的表达增加,这表明 BACH2 抑制了 npTH17 细胞中的 TH17 细胞致病性。

Bach2 KO 对 TH17 细胞的基因表达和染色质可及性有显著的影响。
(c)对 Bach2 KO 和对照 npTH17、pTH17 和 TH1 细胞进行bulk ATAC-seq 分析显示,pTH17 和 npTH17 细胞均偏向 TH1 细胞的染色质谱转变,表明 BACH2 限制了 TH17 细胞中的 TH1 细胞程序。TH1敲除Bach2与否,ATAC-seq的表达差不多,但是pTH17和npTH17在敲除Bach2之后,ATAC-seq的和TH1中交集的峰数量明显增多。尽管 BACH2 缺乏对 TH17 细胞有强烈影响(npTH17 细胞中有 1,118 个 DACR,pTH17 细胞中有 627 个 DACR),但它对 TH1 细胞的染色质谱影响不大(18 个 DACR,FDR < 0.05,| log2(fold change)| > 0.5)。
(b,d)npTH17 和 pTH17 细胞的 Bach2 KO 谱均向 PC1 上的 TH1 细胞偏移,TH1 细胞特异性峰打开,TH17 细胞特异性峰关闭。

因此,pTH17 和 npTH17 细胞中 Bach2 KO 特异性峰附近的基因富集了 TH17 细胞致病性的体内特征,而无致病性和干细胞样表型的特征则被耗尽,这表明 BACH2 调节 TH17 细胞无致病性/干细胞样表型的维持。此外,Bach2 KO 后,npTH17 和 pTH17 细胞中 Treg 和 TH17 细胞的染色质可及性特征均下调。

(上e-g)为了在体内测试 BACH2 在 TH17 细胞中的功能,研究使用了 2D2 EAE 转移模型,其中髓鞘少突胶质细胞糖蛋白 (MOG) 特异性 CD4+ T 细胞 (2D2) 被过继转移。BACH2 缺陷型 2D2 细胞比用病毒处理的对照的 T 细胞诱发了更严重的 EAE 疾病 ,并且在中枢神经系统中 IL-17A+ 和 IL-17A+ IFNγ+ 2D2 细胞的比例明显较高(f),而受体小鼠脾脏中 IL-10+ 2D2 细胞的比例较低(g)。
(下e-f)2D2 细胞在中枢神经系统中的活力和数量没有差异,受体小鼠脾脏和中枢神经系统中 2D2 细胞频率也没有差异。
这些结果证实了 BACH2 是体内 TH17 细胞致病性的负调控因子。
9.BACH2 表达增加可改善 TH17 细胞的自身免疫

(左a)对于 Bach2 过表达 (OE) 分析,研究用含有组成型 Bach2 cDNA 表达盒 (Bach2 OE) 或空对照的逆转录病毒载体转导 pTH17 细胞,然后进行bulk ATAC-seq。
(右a,b)Bach2 OE 导致 pTH17 细胞染色质景观发生重大变化,打开了 524 个峰,如靠近与干细胞样 npTH17 细胞表型 (Ccr7、Maf 和 Tcf7) 相关的基因的峰;关闭了更多峰 (3,438 个峰),如与 TH17 细胞致病性有关的基因的峰,例如 Bhlhe40、Csf2、Cxcr6、Ermn、Ifng 和 Rbpj。
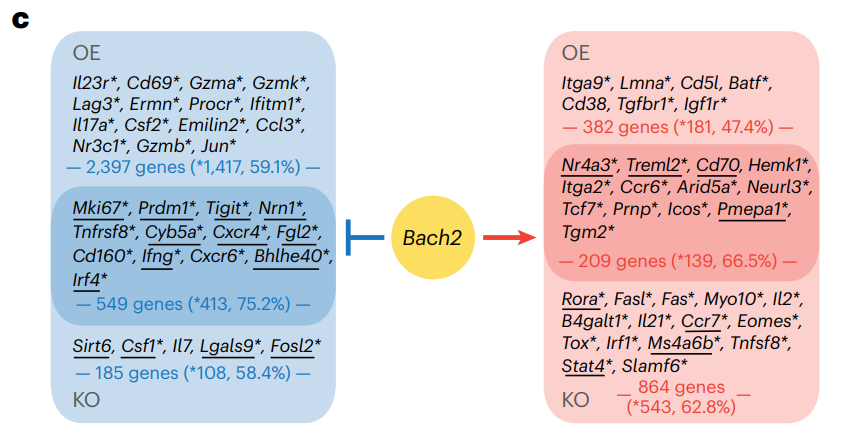
Bach2 KO 和 Bach2 OE 极化的 TH17 细胞中染色质可及性的变化截然相反。图c展示受BACH2调控的关键基因,左边蓝色是被BACH2抑制的基因,右边红色是被BACH2激活的基因。每块被划分为三个小块,从上到下是过表达组(OE)、对照组和敲除组(KO)。星号表示在BACH2 ChIP-seq数据中至少有一个峰与该基因相关。加粗并带下划线的基因表示与Bach2 OE(或KO)相关的峰中具有BACH2基序的基因。
-
蓝色区域(BACH2抑制):上半部分列出了Bach2 OE实验中DACR的2397个基因,其中1417个基因(59.1%)有显著差异;中间部分列出了549个基因,其中413个基因(75.2%)在BACH2 ChIP-seq中有显著峰值;下半部分列出了185个基因,其中108个基因(58.4%)在KO实验中有显著差异。
-
红色区域(BACH2激活):上半部分列出了Bach2 OE实验中DACR的382个基因,其中181个基因(47.4%)有显著差异;中间部分列出了209个基因,其中139个基因(66.5%)在BACH2 ChIP-seq中有显著峰值;下半部分列出了864个基因,其中543个基因(62.8%)在KO实验中有显著差异。
这些 DACR 中的许多都含有 BACH2 结合motif,包括干细胞样 T 细胞标记物 Tcf7 和 Slamf6以及促炎基因 Ifng 和 Csf2 的位点。在 Bach2 KO 中,734 个变得更加开放的基因峰中有 483 个(65.8%),1,073 个变得更加封闭的基因峰中有 208 个(19.4%)含有 BACH2 结合motif。在 Bach2 OE 中,2,946 个变得更加封闭的基因峰中有 1,326 个(45.0%)检测到 BACH2 结合motif,591 个变得更加开放的基因峰中有 126 个(21.3%)检测到 BACH2 结合motif。
此外,在分析中发现的 47.4–75.2% 的 BACH2 靶基因与 BACH2 结合,这是在已发表的诱导 Treg 细胞的 BACH2 染色质免疫沉淀与测序 (ChIP-seq) 数据中发现的,结果表明 BACH2 会与靶基因直接结合。
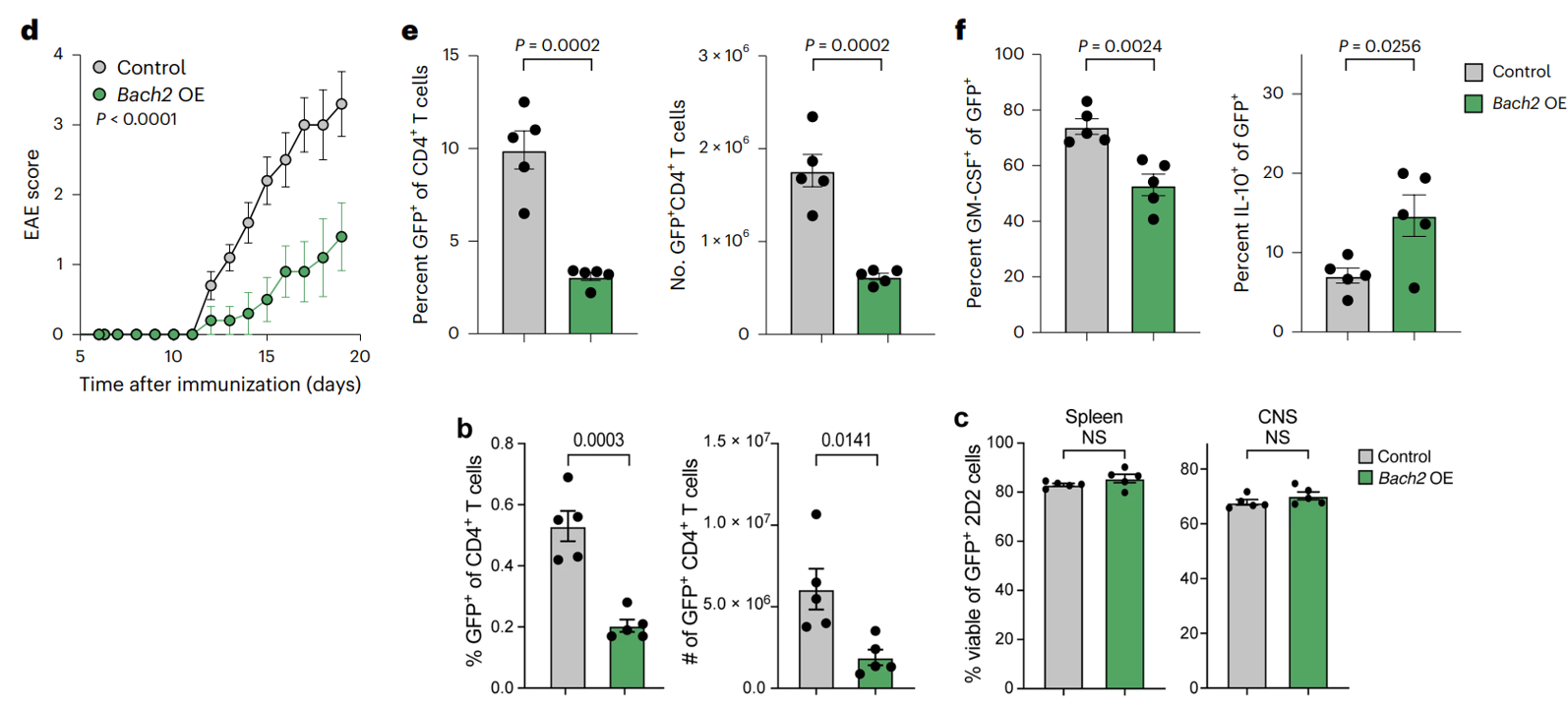
2D2 T 细胞会诱发严重的 EAE 疾病,而过表达 Bach2 的 T 细胞会明显缓和疾病(d);中枢神经系统 (e) 和脾脏 (b) 中转导 (GFP+) T 细胞的比例和数量明显较低,但中枢神经系统或脾脏中的细胞活力没有差异(c)。中枢神经系统中的细胞表达致病性细胞因子 GM-CSF 的比例在过表达Bach2的组中明显降低,而表达调节性细胞因子 IL-10 的比例较高(f)。
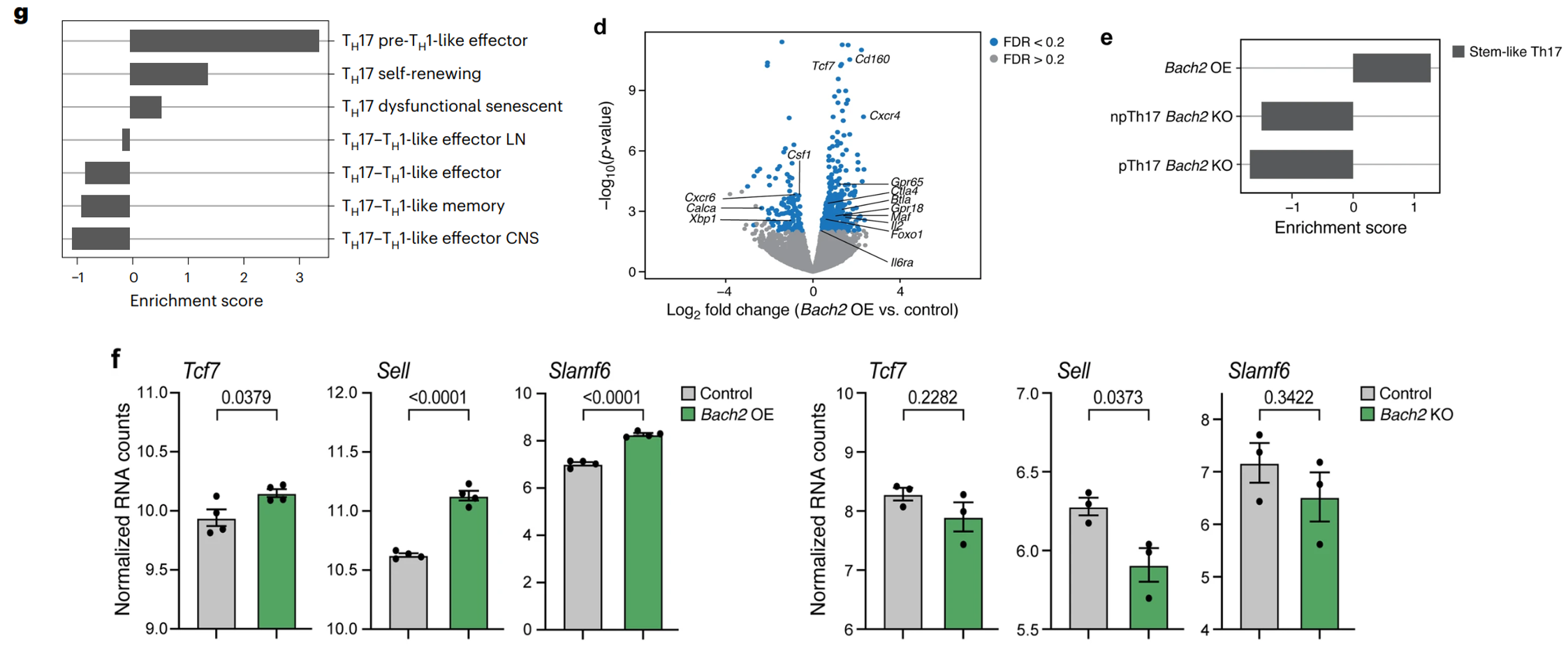
(g,d)EAE 患病小鼠中枢神经系统中的 Bach2 OE 转导(GFP+)细胞的基因表达富集体内无致病性和干细胞样 TH17 细胞的特征,而对照细胞具有高度致病性特征,具有 TH17、前 TH1 效应样细胞的特征。
(e)体内干细胞样 TH17 特征在 Bach2 OE 细胞中富集,而在 Bach2 KO 细胞中下调。
(f)干细胞样基因 Tcf7、Slamf6 和 Sell 在 Bach2 OE 细胞中上调。有研究表明 BACH2 是 CD8+ T 细胞中干细胞样程序的调控因子,结合本研究结果,进一步表明 BACH2 是 T 细胞中干细胞样表达程序的更广泛驱动因素。

为了进一步探索 BACH2 调节 TH17 细胞的潜在机制,研究对 BACH2 为中心进行子网络分析。在该模型中,BACH2 直接激活与 npTH17 细胞相关的 TF 基因(Ahr 和 Rbpj)和基因(Lgals1、Vim 和 Ctsw),并抑制与 pTH17 和 TH1 细胞相关的 TF 基因(Id3 和 Tbx21)和基因(Ccl5、Ifit1、Ms4a4b、Ccl3 和 Ccl4)。此外,分析揭示了 BACH2 与 TF RBPJ 之间的密切关系,其中 BACH2 直接激活 RBPJ 表达,并且 BACH2 和 RBPJ 有多个相同直接靶基因,表明它们以共同作用以激活 npTH17 程序并抑制 pTH17 程序。
总的来说,这些结果验证了 BACH2 是体内 TH17 细胞致病染色质可及性的新型调控因子,并表明 BACH2 是治疗自身免疫性疾病的潜在靶点。
10.人类 BACH2 变异与自身免疫性疾病风险相关

(a)BACH2 被列为体外培养的人类 TH17 细胞的top 调控因子之一,表明 BACH2 与人类自身免疫性疾病之间存在潜在关联。
尽管 BACH2 基因座中的遗传变异以前与多种自身免疫性疾病有关,但尚未发现 BACH2 变异与其在 CD4+ T 细胞中的表达和功能之间的关系。
(b-c)图展示了在 rs72928038 上游和下游 1MB 范围内的变体中,每个变体(点)与 GWAS 中的 MS 风险,和作为 BACH2 顺式 eQTL的 MS 风险的关联显著性。该基因座的精细定位报告了三种可解释 GWAS 信号的变体:rs72928038(后验概率:84.2%)、rs10944479(后验概率:10.4%)和 rs6908626(后验概率:1.1%)。
BACH2基因与rs72928038位点的关联性很高,提示BACH2可能通过该位点调控基因表达,进而影响MS的发病风险。
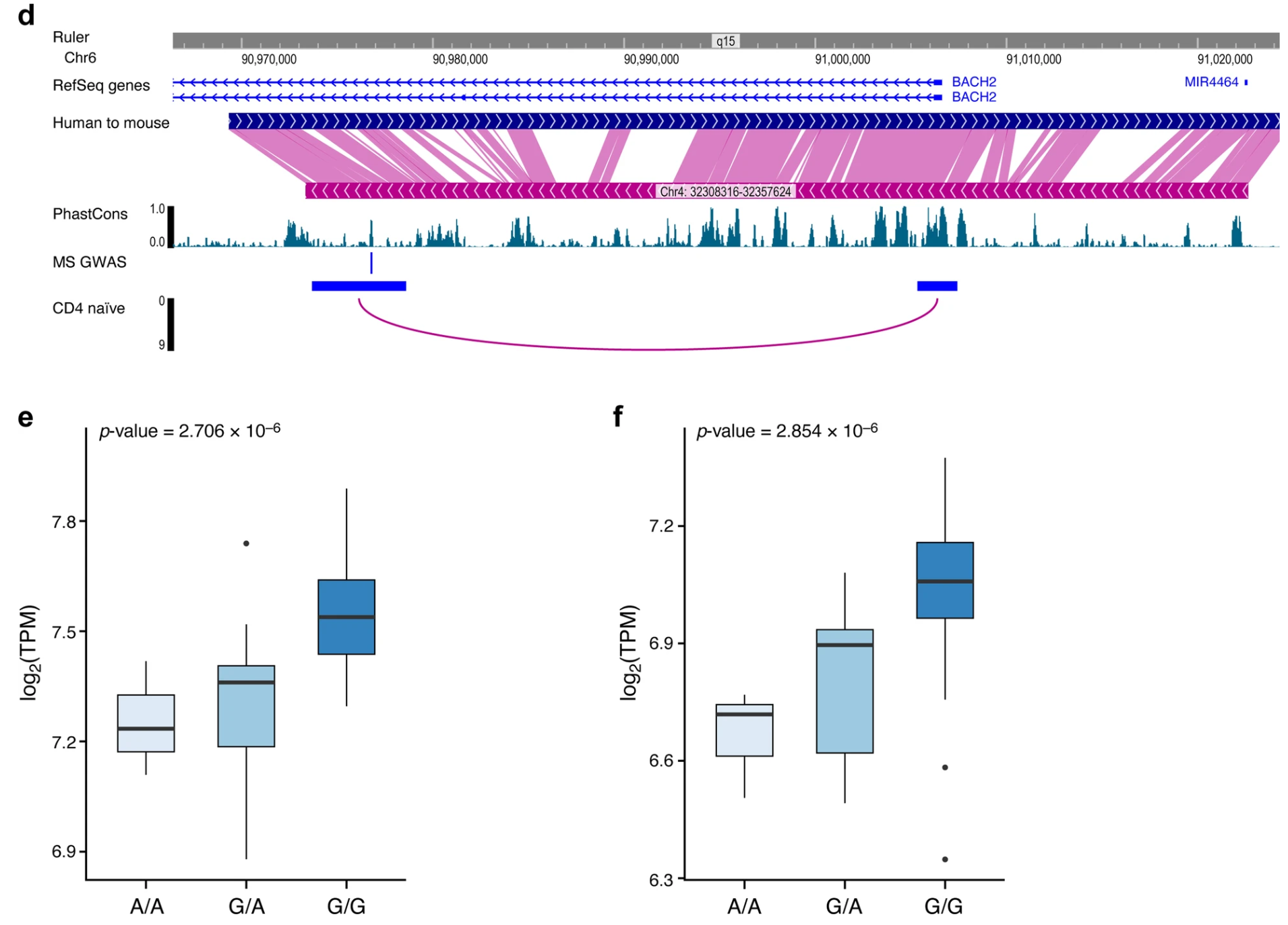
接下来,分析了最强关联的变体 rs72928038 是否可能参与 CD4+ T 细胞中 BACH2 的转录调控。染色质可及性、染色体构象和表达数据表明,变体 rs72928038 位于一个调控元件中,该元件的功能可能与 BACH2 转录调控有关。
(d)首先,rs72928038 位于 BACH2 上一个开放的染色质峰内含子中,该峰在所有原代免疫细胞中都是开放的,表明可能调控附近的基因。此外,启动子捕获相互作用数据显示,在所有 17 种检查的细胞类型和状态下,rs72928038 基因座和 BACH2 启动子之间存在相互作用,但仅在幼稚 CD4+ T 细胞中达到研究范围内的显著性。
(e-f)对公开的 eQTL 数据的分析表明,rs72928038 和 rs10944479仅与 BACH2 基因表达的变化有关。rs72928038 G 等位基因的携带者在幼稚 CD4+ T 细胞和 Treg 细胞中具有更高的 BACH2 表达。
这些结果表明,在多发性硬化症(MS)的易感性中,某些特定的保护性等位基因能够提升CD4+ T细胞中BACH2基因的表达,从而降低MS的发病风险。这与本研究在小鼠 TH17 细胞中的分析结果一致。
总结
研究内容
-
本研究对体外和体内衍生的npTH17和pTH17细胞及其他CD4+ T细胞亚群的染色质景观进行了详细表征。
-
区分npTH17和pTH17细胞在染色质和基因表达水平上稳定的细胞命运与仅基于表达的可变细胞状态。
-
发现基于基因表达研究未识别的调控程序和靶标。
研究方法与优势
综合分析染色质可及性和基因表达,利用多种技术对npTH17和pTH17细胞及其他CD4+ T细胞亚群进行详细表征。通过整合单细胞层面的染色质可及性和基因表达数据,揭示了潜在的调控因子。
结合了染色质可及性和基因表达数据,提供了更全面的细胞状态和命运的理解。揭示了npTH17和pTH17细胞在体内和体外环境中稳定的染色质差异,强调了其稳定性和特异性。识别了如Bach2和Atf3等调控因子,这些因子在仅分析RNA谱的研究中未被发现,展示了本研究方法的独特优势。
研究意义
-
细胞命运研究:通过区分稳定的细胞命运和可变的细胞状态,深化了对npTH17和pTH17细胞生物学特性的理解。
-
调控网络:发现并描述了pTH17细胞和TH1细胞共享的广泛调控网络,确定了pTH17细胞作为npTH17和TH1细胞间的中间细胞命运。
-
BACH2的作用:验证了BACH2在抑制TH17细胞致病性、支持类干细胞TH17细胞程序以及在自身免疫疾病中的重要作用,揭示了BACH2作为潜在治疗靶点的可能性。
-
治疗潜力:BACH2在人体CD4+ T细胞中的表达增加与MS等自身免疫疾病的保护性作用相关,提示诱导BACH2表达和/或活性可能成为抑制致病效应T细胞的有效治疗策略。