2023年10月24日,西北农林科技大学园艺学院管清美教授团队在Plant Biotechnology Journal(最新IF:13.8)上发表题为“The chromatin remodeller MdRAD5B enhances drought tolerance by coupling MdLHP1-mediated H3K27me3 in apple”的研究性论文。研究报告了一种干旱诱导的染色质重塑因子MdRAD5B,通过调节干旱响应基因上的染色质的可及性和H3K27me3沉积来增强苹果的耐旱性。研究结果为苹果响应干旱胁迫的机制提供了新的见解,并为耐旱作物改良提供了候选基因。研究所用ATAC-seq、RNA-seq和H3K27me3 ChIP-seq技术手段爱基百客均可提供。

01 研究背景
苹果是世界范围内温带地区最重要的经济水果作物之一,但在气候变化的背景下,苹果树常遭受干旱胁迫的挑战。分子育种是增强苹果抗旱性的有效方法,了解苹果应对干旱胁迫的潜在机制至关重要。尽管已经研究了苹果响应干旱的生理变化、转录调节和表观遗传调节,但是参与响应干旱的染色质重塑仍然难以捉摸。
02 研究路线

03 研究结果
-
1、MdRAD5B正调节耐旱
干旱胁迫处理能显著诱导金冠苹果中MdRAD5B的表达(图1A),而器官特异性表达分析显示其主要在根和叶中表达(图1B)。MdRAD5B定位于细胞核(图1C),从绿藻到木兰植物中序列保守(图1D)。这都表明MdRAD5B是一种潜在的干旱调节因子。瞬时沉默MdRAD5B(RNAi)的植物(图2A),存活率降低(图2B),电解质渗漏率和丙二醛(MDA)含量增加(图2C, D);过表达植物则表现相反(图2E-H)。这些结果表明MdRAD5B是苹果抗旱性的正向调节因子。
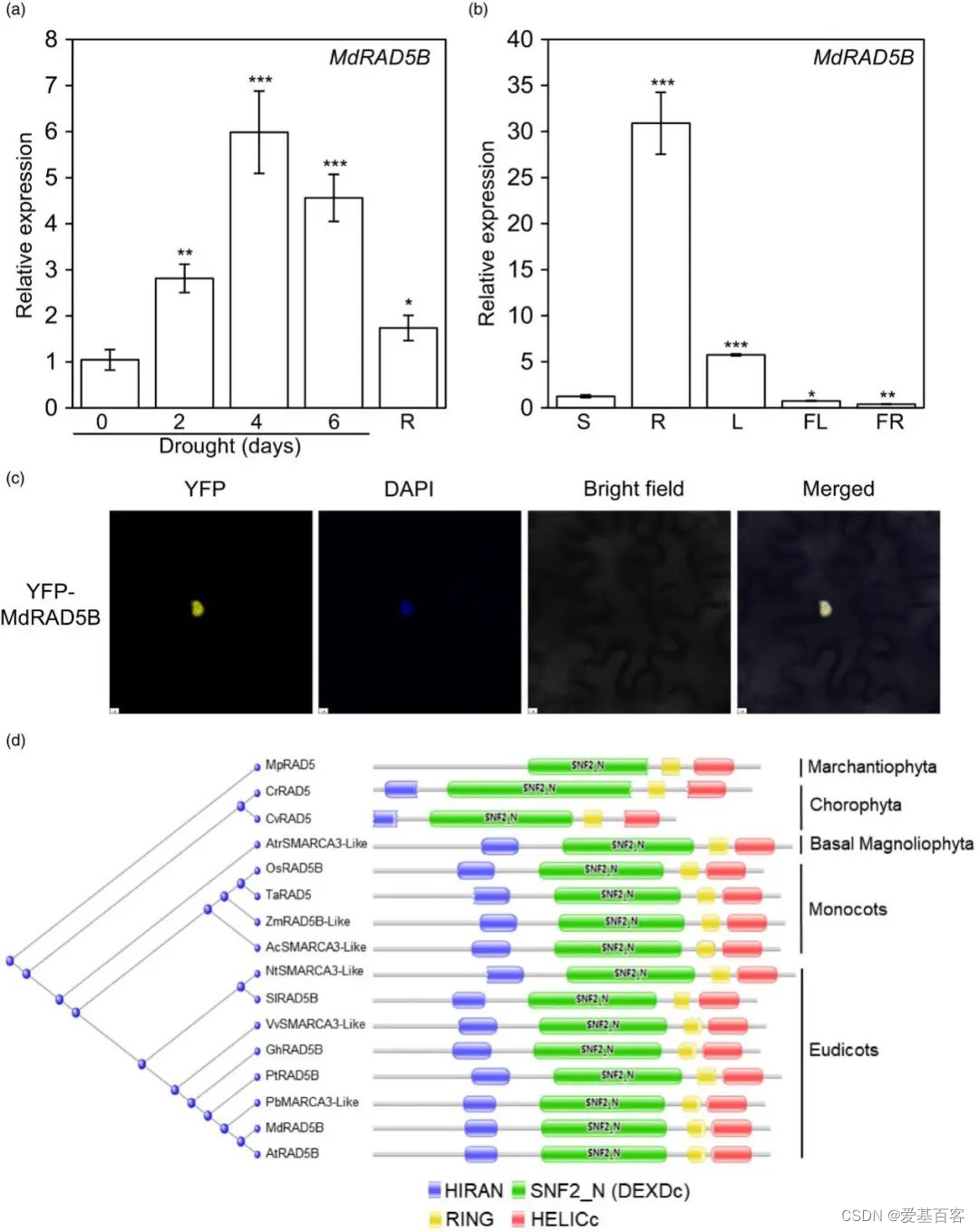
图1. MdRAD5B响应干旱胁迫并定位于细胞核

图2. MdRAD5B在耐旱性中发挥正向作用
-
2、MdRAD5B调控干旱响应基因的表达
对脱水条件下的MdRAD5B RNAi和GL-3植物进行RNA-seq分析,鉴定了1,997个干旱抑制基因在脱水后的RNAi植物中上调,而945个干旱诱导基因在脱水后下调(图3A-C)。10个已知的与耐旱性正相关或负相关的基因被选择来做qRT-PCR,证明了RNA-seq的结果并表明MdRAD5B调控干旱响应基因(图3D-M)。
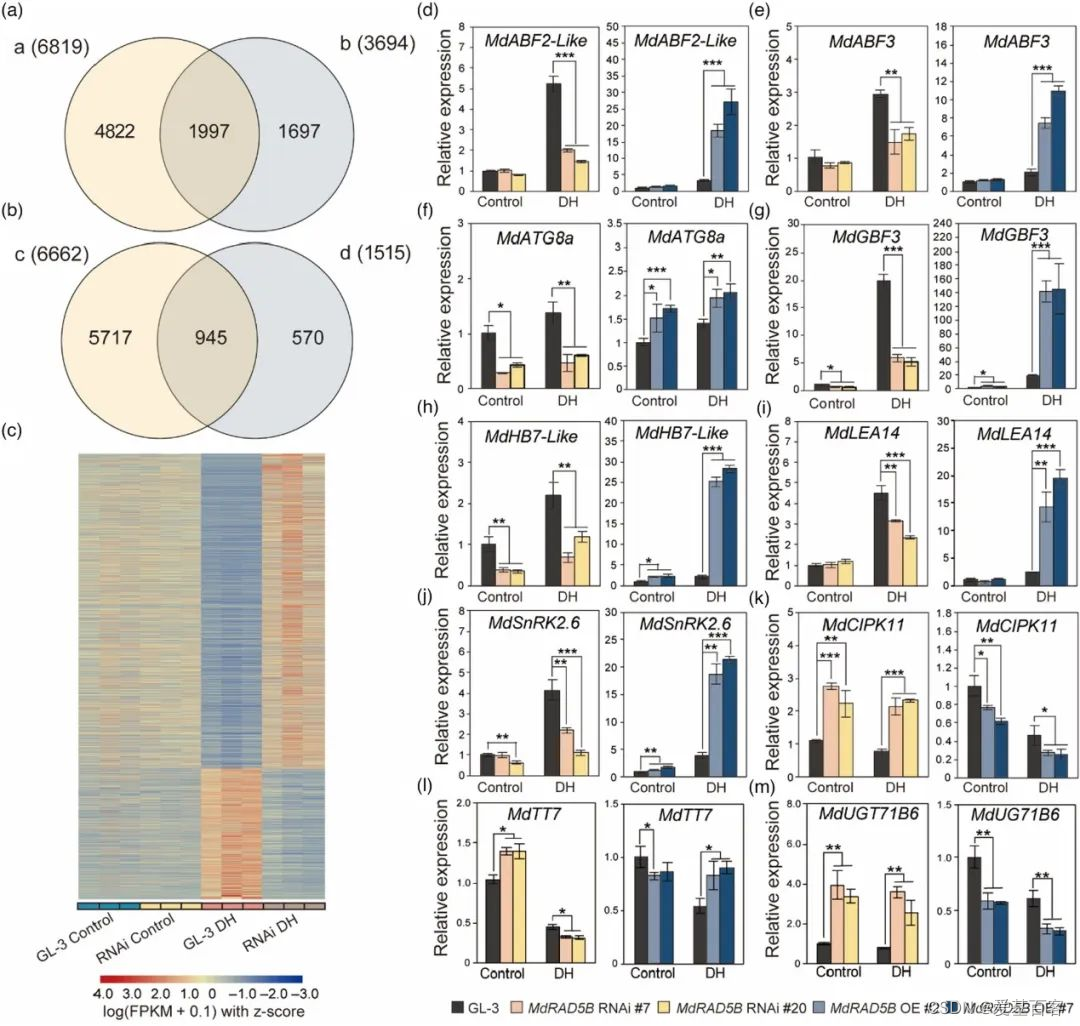
图3. MdRAD5B在响应缺水中调节干旱响应基因
-
3、MdRAD5B调控干旱响应基因的染色质重塑
体外酶活测定表明DNA可以刺激含有DEXDc结构域的MdRAD5B重组蛋白的ATP酶活性(图4A),而当DNA缺失或者MdRAD5B的K562(ATP酶活性的关键位点)突变为A562后,活性消失,暗示MdRAD5B是一种潜在的染色质重塑因子。利用ATAC-seq对正常和缺水条件下的RNAi个GL-3植物进行检测,GL-3在缺水处理前后有17,010个差异可及性区域(DARs),而正常和缺水条件下RNAi和GL-3之间分别有6,436和7,369个DARs,且主要在启动子区富集(图4B),说明MdRAD5B可以调节干旱胁迫下染色质的可及性。7,369个DARs被进一步注释到5,815个基因,它们参与激素应答、非生物胁迫和发育过程(图4C)。整合ATAC-seq和RNA-seq,发现与GL-3相比,缺水条件下的RNAi中有220个基因表达和可及性均上调,246个基因表达和可及性都下调(图4D, E),其中不乏一些已知的正调控耐旱性因子(如MdRCI2A、MdNF-YB3、MdXERICO、MdGBF3和MdABF2-Like)和负调控耐旱性因子(如MdGIM2)。缺失条件下,与GL-3相比,RNAi植物中MdRCI2A和MdNF-YB3表达和可及性都降低(图4F),而MdGIM2的表达和可及性都增强。这些数据表明,MdRAD5B至少在一定程度上通过调节染色质可及性来调节干旱响应基因的表达。

图4. MdRAD5B在响应缺水中调节染色质可及性
-
4、MdRAD5B与MdLHP1相互作用并通过不依赖泛素的20S蛋白酶体介导其降解
利用酵母筛库,研究获得了一些与MdRAD5B相互作用的蛋白,包括拟南芥中负调控耐旱性的H3K27me3阅读器LHP1的同源物。酵母双杂交(Y2H)、分裂-荧光素酶互补(Split-LUC)和免疫共沉淀(Co-IP)实验进一步证实了MdRAD5B和MdLHP1在体内的相互作用(图5A-E)。Y2H的结果还表明MdRAD5B通过RING结构域与MdLHP1互作,这种结构域通常参与蛋白质降解。在烟草叶片中共表达Myc-MdLHP1和MdRAD5B- flag时,Myc-MdLHP1的量大幅减少(图5F),表明MdRAD5B对MdLHP1进行了降解。此外,与GL-3相比,正常和缺水条件下,MdRAD5B-OE和MdRAD5B-RNAi植株中的MdLHP1蛋白水平分别降低和升高(图5G, H),进一步表明MdRAD5B在干旱条件下降解MdLHP1。在共表达实验中添加蛋白酶抑制剂MG132后,MdRAD5B介导的MdLHP1降解被抑制,这引起了作者的注意。作者分别进行了以泛素化的MdBBX7作为阳性对照,以及添加或不添加PYR-41(选择性地抑制泛素依赖的26S蛋白酶体途径,而不抑制泛素不依赖的20S蛋白酶体途径)的实验(图5I,J),结果表明MdRAD5B主要以泛素不依赖的20S核心蛋白酶体降解MdLHP1。进一步的实验证实,20S核心颗粒蛋白酶体(CP)的β1亚基(MdPBA1)是介导MdRAD5B降解MdLHP1的核心组分(图5K)。

图5. MdRAD5B通过不依赖泛素的20S蛋白酶体与MdLHP1相互作用并降解
-
5、MdRAD5B调节干旱反应基因上的H3K27me3沉积以响应干旱
AtLHP1可以与MSI1相互作用募集PRC2复合体维持H3K27me3组蛋白修饰,作为其同源物MdLHP1也可以结合H3K27me3并维持H3K27me3的水平。正常和缺水条件下,MdRAD5B-OE植物里的H3K27me3水平高于GL-3,RNAi的植物表现相反(图6A, B)。作者怀疑MdRAD5B是否也会影响干旱胁迫下H3K27me3的沉积,因此进行了ChIP-seq实验。正常和缺水条件下,RNAi和GL-3植物之间分别有2,244和633个差异H3K27me3修饰峰。其中,633个差异H3K27me3修饰峰主要位于启动子区,与615个基因相关,这些基因主要富集在代谢过程以及对非生物应激和激素的响应中(图6D)。此外,与GL-3相比,RNAi植株在缺水条件下的DEGs中有66个差异甲基化基因(图6E)。选择6个与耐旱性相关的基因进行ChIP-qPCR和qRT-PCR验证。在缺水条件下,与GL-3相比,RNAi植株中4个超甲基化基因的H3K27me3沉积增加,表达降低;2个低甲基化基因的H3K27me3沉积减少,表达增加(图6F-H, 3L, M)。这些结果表明,MdRAD5B可以通过调控干旱胁迫下H3K27me3的沉积来调控干旱响应基因的表达。

图6. MdRAD5B调节H3K27me3沉积以响应脱水
-
6、MdRAD5B在MdLHP1的上游起作用
遗传分析显示,PEG处理14d后,MdLHP1抗愈伤组织的相对生长率高于野生型,而MdRAD5B-RNAi愈伤组织的相对生长率低于野生型(图7A, B)。此外,减少MdLHP1的表达可以恢复PEG处理下MdRAD5B-RNAi愈伤组织相对生长率的下降,表明在干旱胁迫响应中,MdLHP1作用于MdRAD5B的下游。此外,PEG处理下,与野生型相比,MdTT7、MdNHL13和MdAHSP等干旱正 调节基因在MdRAD5B-RNAi愈伤组织中H3K27me3的富集程度更高而表达更低,然而MdLHP1抗愈伤组织中这些基因的H3K27me3的富集程度较低而表达更高,且MdLHP1表达降低会阻碍MdRAD5B-RNAi愈伤组织中这些基因的H3K27me3的富集程度升高并恢复表达(图7C, D)。干旱负调节基因MdIAA16在转基因愈伤组织里的H3K27me3和表达趋势与正响应基因相反。这些结果进一步表明,MdRAD5B在MdLHP1上游发挥作用,至少在一定程度上通过破坏干旱胁迫下MdLHP1的稳定性,调节干旱响应基因(包括MdTT7、MdNHL13、MdAHSP和MdIAA16)的H3K27me3沉积和表达。

图7. MdRAD5B和MdLHP1的遗传分析
-
8、RING和DEXDc结构域是MdRAD5B介导耐旱性所必需的
DEXDc结构域和RING结构域分别负责MdRAD5B的ATP酶活和MdLHP1的互作,研究生成了MdRAD5B-OE、MdRAD5B-ΔRING-OE(缺失RING结构域)和MdRAD5B-ΔDEXDc-OE(缺失DEXDc结构域)转基因植物。PEG处理后14d,MdRAD5B-OE愈伤组织的相对生长率高于野生型,而MdRAD5B-ΔRING-OE和MdRAD5B-ΔDEXDc-OE的相对生长率低于MdRAD5B-OE愈伤组织,但高于野生型。这些数据表明,MdRAD5B在苹果耐旱性中的功能需要DEXDc和RING结构域。RAD5A的HIRAN结构域负责植物DNA修复,而RAD5B的HIRAN结构域可能具有不同的功能。研究还生成了MdRAD5B-ΔHIRAN -OE(缺失HIRAN结构域)的愈伤组织,PEG处理后,其相对生长率与MdRAD5B-OE愈伤组织的相当,尽管它大于野生型,表明MdRAD5B的HIRAN结构域在苹果耐旱性中不是必需的。
04 研究结果
综上所述,这项研究证明MdRAD5B通过其染色质重塑功能或通过调节H3K27me3来调节干旱响应基因的表达水平,以应对干旱胁迫(图8)。研究工作不仅为染色质重塑因子在植物干旱响应中的重要性提供了新的证据,而且也有利于未来对水果作物的基因工程。
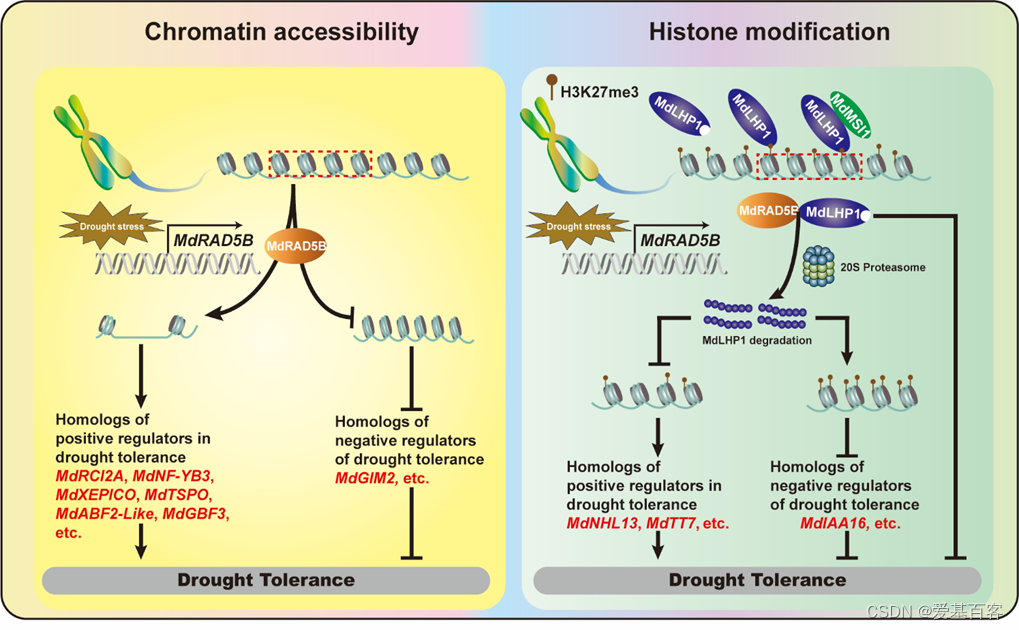
图8. MdRAD5B响应干旱胁迫的工作模型
# 关于我们 #
爱基百客专注于提供领先表观组学服务。公司先后引入ChIP、CUT&Tag、WGBS、ATAC-seq、全转录组、10x Genomics、DNBSEQ-T7等实验平台,不断提升公司的科研服务能力。至今合作的科研客户超2000家,涵盖国内知名科研院所、高校以及相关生物企业,科研成果曾多次在Cancer Cell、Nature Communications、J HEMATOL ONCOL、Plant Cell 等国际高水平学术期刊发表,受到了客户广泛好评,是国内成长最迅速的高通量测序科研服务企业之一。